Навигатор Garmin Gpsmap 196 010-00301-51
Летайте свободно вместе с авиационным навигатором GPSMAP 196. Универсальный монохромный авианавигатор для работы в воздухе, на воде и на суше.
Три стихии в ваших руках?
Навигатор умеет работать в трех режимах – на суше, на море и в воздухе. Встроенная базовая карта с функцией автоматического расчета маршрутов позволяет без труда менять один вид транспорта на другой, а также путешествовать пешком. Кроме того, вы можете установить в аппарат морскую картографию.
«Модель GPSMAP 196 содержит функцию WAAS, подробные карты, систему рулевого управления HIS, базу данных Jeppesen, а также другие специальные функции для пилотов. Для рулевого управления используется страница HIS с индикатором VNAV, благодаря чему Вы будете предупреждены об опасных ситуациях»Сотрудник фирменного магазина Garmin
Весь мир на ладони
Навигатор отличается большим контрастным цветным дисплеем, на котором информация карты видна до мельчайших деталей.
Следите за полетом по приборной панели
Навигатор оснащен уникальной функцией – страницей, имитирующей приборную панель самолета для графической индикации информации о полетах.
Обстановка под контролем
Даже без загруженных GPS-карт навигатор имеет доступ к базе Jeppesen с указанием аэропортов и других необходимых объектов, а также базами данных рельефа и препятствий на территории США, обеспечивая реалистичную топографию. В режиме рельефа прибор выдает предупреждения типа TAWS при обнаружении препятствий рядом с маршрутом.
Автоматизация журнала полетов
При использовании бесплатного программного обеспечения Garmin FlightBook ведение бортового журнала значительно упрощается – вы можете создавать, управлять, анализировать, сохранять и распечатывать данные полетов из журнала навигатора.
Ремонт Garmin gpsmap 196 в Москве
Причины неисправности навигатора
Сервисный центр Ремфон выполняет качественный ремонт навигатора Garmin gpsmap 196. При этом у насдоставка по России и регулярные акции при минимальных сроках восстановления. Как и любые электронные технически сложные устройства, навигатор gpsmap 196 рано или поздно может выйти из строя. При этом его поломки могут быть, как аппаратными, так и программными. Наши опытные инженеры быстро определяют точную причину неисправности и в минимальные сроки проведут её устранение.
Наиболее распространенные поломки
У навигационных устройств Garmin бывают следующие неисправности:
- слабый сигнал спутников
- садится аккумулятор и батарейки хватает на несколько часов
- трещины на задней панели
Большая часть повреждений GPS-навигаторов связана с неосторожным обращением, поэтому часто
требуется Замена клавиши громкости.
Наш центр выбирают многие из тех, кому нужно отремонтировать навигатор gpsmap 196, так, как только у нас:
- честные цены
- приятные цены
- полная диагностика
- гарантия от 3 месяцев
- положительные отзывы о сервисе
- {advantages}
А, если вам необходимо уточнить некоторые вопросы по данной услуге, вы всегда можете связаться с нашими специалистами по телефону+7 (495) 699-14-10 или в паблике Фейсбука.
Бесплатная диагностика
даже при отказе от ремонта
Оригинальные запчасти
для ремонта. С гарантией.
Срочный ремонт
до 30 мин
Гарантия на все выполненные
работы и установленные запчасти
Рейтинг на yell. ru:
ru:
Рейтинг yandex.ru:
Отзывы:
5/5 Отзывов: 17
| 80% ремонтов делаются в тот же день | цена | стоимость детали включена |
|---|---|---|
| Диагностика | 0 | |
| Прошивка (обновление ПО) | 1000 | |
| Понижение (откат) версии ПО | 2000 | |
| Русификация (локализация) | ||
| Замена динамика | 2400 | |
| Замена аккумулятора (АКБ) | 900 | |
| Замена дисплея (стекла) | 1100 | |
| Замена тачскрина (сенсора) | 1100 | |
| Замена задней крышки | 800 | |
| Замена элементов корпуса | 1200 | |
| Замена системной (материнской) платы | 1000 | |
| Замена разъема питания (подзарядки) | 2000 | |
| Замена разъема внешней антенны | 1100 | |
| Замена кнопки включения | 2500 | |
| Замена клавиши громкости | 1200 | |
| Замена разъема флеш карты | 4000 | |
| Замена микросхемы питания | 1600 | |
| Замена контроллера зарядки | 1600 | |
| Замена GPS модуля | 1600 | |
| Замена GPS антенны | 1500 | |
| Восстановление после попадания воды | 1900 | |
| Восстановление данных | 1500 |

 org/OfferCatalog»>
org/OfferCatalog»> org/OfferCatalog»>
org/OfferCatalog»> org/OfferCatalog»>
org/OfferCatalog»> org/OfferCatalog»>
org/OfferCatalog»> org/OfferCatalog»>
org/OfferCatalog»> org/OfferCatalog»>
org/OfferCatalog»> org/OfferCatalog»>
org/OfferCatalog»> org/OfferCatalog»>
org/OfferCatalog»> org/OfferCatalog»>
org/OfferCatalog»> org/OfferCatalog»>
org/OfferCatalog»>Вопросы и ответы по ремонту Garmin gpsmap 196
Добрый день, у меня навигатор Garmin gpsmap 196 сгорел при зарядке от бортовой сети, возможен ли ремонт и сколько будет стоить?
Здравствуйте Гафур, ремонт Гармина возможен, стоить он будет начиная от 3000р.
Мы также ремонтируем
OZON.ru
Пермь- Ozon для бизнеса
- Мобильное приложение
- Реферальная программа
- Зарабатывай с Ozon
- Подарочные сертификаты
- Помощь
- Пункты выдачи
- TOP Fashion
- Premium
- Ozon Travel
- Ozon Счёт
- LIVE
- Акции
- Бренды
- Магазины
- Сертификаты
- Электроника
- Одежда и обувь
- Детские товары
- Дом и сад
- Зона лучших цен
Такой страницы не существует
Вернуться на главную Зарабатывайте с OzonВаши товары на OzonРеферальная программаУстановите постамат Ozon BoxОткройте пункт выдачи OzonСтать Поставщиком OzonЧто продавать на OzonSelling on OzonО компанииОб Ozon / About OzonВакансииКонтакты для прессыРеквизитыАрт-проект Ozon BallonБренд OzonГорячая линия комплаенсУстойчивое развитиеOzon ЗаботаПомощьКак сделать заказДоставкаОплатаКонтактыБезопасностьOzon для бизнесаДобавить компаниюМои компанииПодарочные сертификаты © 1998 – 2022 ООО «Интернет Решения». Все права защищены.
Версия для слабовидящихOzonИнтернет-магазинOzon ВакансииРабота в OzonOZON TravelАвиабилетыRoute 256Бесплатные IT курсыLITRES.ruЭлектронные книги
Все права защищены.
Версия для слабовидящихOzonИнтернет-магазинOzon ВакансииРабота в OzonOZON TravelАвиабилетыRoute 256Бесплатные IT курсыLITRES.ruЭлектронные книгиГармин подавляет пролиферацию и миграцию клеток рака яичников человека посредством ингибирования пути ERK/CREB
Введение
Рак яичников является шестой по частоте причиной
смертность от рака среди западных женщин. Хотя он представляет
30% случаев рака женских половых органов приходится на рак яичников.
ответственны за более чем половину смертей (1). Значительный прогресс был достигнут в
хирургия и совершенствование химиотерапевтических средств при яичниках
рак, что привело к улучшению 5-летней выживаемости, однако
Рак остается ведущей причиной смерти всех гинекологических
рак (1,2).Основная задача нынешних
химиотерапия для лечения рака яичников связана с лекарственными средствами
сопротивление или рецидив. Таким образом, идентификация нового противоракового
лекарства и понимание лежащих в их основе молекулярных механизмов. большую помощь в лечении рака яичников.
большую помощь в лечении рака яичников.
Натуральные продукты привлекли значительное
внимание из-за их мощного противоракового действия в прошлом
десятилетия (3,4). Подсчитано, что более 46%
новые лекарства и новые лекарства-кандидаты для лечения рака, одобренные США
Управление по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA) в период с 1989 по 1995 г.
натуральные продукты или производные натуральных продуктов (5).Примечательно, что недавние исследования
показали, что β-карболиновые алкалоиды, большая группа
природные и синтетические индольные алкалоиды, широко распространенные в
экстракты из листьев, коры, корней и семян различных
растений, обладают широким спектром психофармакологического и
нейрофармакологические эффекты (6).
Многие алкалоиды нашли широкое применение в клинической практике.
кашель, гипертония и воспаление. Среди них гармин
(7-метокси-1-метил-9Hпиридо[3,4-b]индол), который первоначально
выделен из семян гармала обыкновенного и
Banisteriopsis caapi и традиционно использовался в народных целях. медицина на Ближнем Востоке, в Центральной Азии и Южной Америке
(7), по-видимому, проявляет различные
фармакологические эффекты как in vitro, так и in vivo,
такие как антиальцгеймеровские, противовоспалительные анксиолитические и
антидепрессивные эффекты (8).Примечательно, что недавние исследования также показывают, что гармин оказывает
значительный противораковый эффект в некоторых раковых клетках, включая
клетки гепатобластомы HepG2 (9),
клетки меланомы B16F-10 (10) и
клетки рака желудка MGC-803 (11).
Однако влияние гармина на пролиферацию и миграцию
клеток рака яичников остается неизвестным.
медицина на Ближнем Востоке, в Центральной Азии и Южной Америке
(7), по-видимому, проявляет различные
фармакологические эффекты как in vitro, так и in vivo,
такие как антиальцгеймеровские, противовоспалительные анксиолитические и
антидепрессивные эффекты (8).Примечательно, что недавние исследования также показывают, что гармин оказывает
значительный противораковый эффект в некоторых раковых клетках, включая
клетки гепатобластомы HepG2 (9),
клетки меланомы B16F-10 (10) и
клетки рака желудка MGC-803 (11).
Однако влияние гармина на пролиферацию и миграцию
клеток рака яичников остается неизвестным.
С этой целью настоящее исследование было направлено на
исследовать противораковый эффект и возможные механизмы
опосредованная гармином пролиферация и миграция клеток в SKOV-3 человека
клетки.Наши результаты показали, что гармин значительно
подавляет пролиферацию и миграцию клеток рака яичников
через ингибирование пути ERK/CREB. Более того, уменьшенный
экспрессия фактора роста эндотелия сосудов (VEGF), матрикс
металлопротеиназы (ММП)-2 и ММП-9 могут участвовать в
гармин-опосредованная пролиферация клеток SKOV-3.
Материалы и методы
Антитела и реагенты
Антитела к фосфо-ERK1/2 (Thr202/Tyr204) (кат. № 9101), ERK1/2 (кат.нет. 9102), фосфо-CREB (Ser133) (антитела к фосфо-ERK1/2 (Thr202/Tyr204) (кат. № 9198), CREB (антитела к фосфо-ERK1/2 (Thr202/Tyr204) (каталожный номер 9197), β-актин (антитела к фосфо-ERK1/2 (Thr202/Tyr204) (кат. № 4970), козий антикроличий пероксидаза хрена (HRP), связанный антитело (антитела к фосфо-ERK1/2 (Thr202/Tyr204) (каталожный номер 7074), а также U0126 (специфический ERK1/2 ингибитор) были приобретены у Cell Signaling Technology (Beverly, Массачусетс, США). Гармин, эпидермальный фактор роста (EGF), диметил сульфоксид (ДМСО) и 3-(4,5-диметилтиазол-2-ил)-2,5-дифенил бромид тетразолия (MTT) были приобретены у Sigma-Aldrich (St.Луис, Миссури, США). Культуральная среда и другие растворы, используемые для клеток культуры были приобретены у Invitrogen (Шанхай, Китай). SYBR® Premix Ex Taq™ II был получен от Takara Bio, Inc. (Оцу, Япония).
Культура клеток и обработка
клеток рака яичников человека SKOV-3 выращивали в
Среда RPMI-1640 с добавлением 10% эмбриональной бычьей сыворотки, 50 мкг/мл
стрептомицин, пенициллин 50 МЕ/мл. Культуры поддерживали при 37°C.
во влажной атмосфере, содержащей 5% CO2. Клетки были
регенерировали каждые 3 дня при достижении 70–90% слияния.Гармин растворяли в ДМСО и разбавляли до подходящей концентрации.
концентрации с питательной средой. Конечная концентрация ДМСО
в культуральной среде не превышала 0,1%.
Культуры поддерживали при 37°C.
во влажной атмосфере, содержащей 5% CO2. Клетки были
регенерировали каждые 3 дня при достижении 70–90% слияния.Гармин растворяли в ДМСО и разбавляли до подходящей концентрации.
концентрации с питательной средой. Конечная концентрация ДМСО
в культуральной среде не превышала 0,1%.
ПЦР с обратной транскрипцией
Тотальную РНК экстрагировали с использованием реагента TRizol.
Обратную транскрипцию проводили согласно инструкции производителя.
инструкции от Invitrogen. ПЦР-анализ проводили с использованием
следующие смысловые и антисмысловые праймеры: Праймеры VEGF были
прямой 5′-CTGGGCTGTTCTCGCTTCG-3′ и обратный
5′-CTCTCCTCTTCCTTCTCTTCTTCC-3′.Праймеры ММП-2 были
прямой 5′-CCGTCGCCCATCATCAAGTTC-3′ и обратный
5′-GCAGCCATAGAAGGTGTTCAGG-3′. Капсюли ММП-9 были выдвинуты вперед
5′-TGGTCCTGGTGCTCCTGGTG-3′ и обратный 5′-GCTGCCTGTCGGTGAGATTGG-3′.
Праймеры GAPDH были прямыми 5′-ACAACTTTGGTATCGTGGAAGG-3′.
и обратный 5′-GCCATCACGCCACAGTTTC-3′. Этапы амплификации были
следующим образом: 94°С в течение 5 мин, 30 циклов денатурации в течение 45 с при
95°С, отжиг 45 сек при 60°С, элонгация при 72°С 45 сек,
и удлинение при 72°С в течение 1 мин. После амплификации ПЦР
продукты подвергали электрофорезу на 1.5% агарозный гель и визуализировали
окрашивание бромистым этидием.
После амплификации ПЦР
продукты подвергали электрофорезу на 1.5% агарозный гель и визуализировали
окрашивание бромистым этидием.
Количественная ПЦР в реальном времени (кОТ-ПЦР)
кДНК получали, как описано выше. Используемые праймеры для ПЦР для анализа перечислены выше. Система ПЦР на 20 мкл была состоит из 2 мкл кДНК, 10 мкл SYBR Premix Ex Taq II, 0,4 мкл ROX Эталонный краситель II, 0,8 мкл 10 мкМ прямой праймер, 0,8 мкл 10 мкМ обратный праймер и 6 мкл ddh3O. Амплификация qRT-PCR была выполняется при следующих условиях: 1 цикл начального денатурация 30 сек при 95°С, 40 циклов денатурации 5 сек при 95°С с последующим синтезом ДНК в течение 30 сек при 60°С.После усиление, сбор и анализ данных в режиме реального времени. выполненный.
Вестерн-блот анализ
После обработки различными соединениями клетки
собирают в указанное время, промывают и подвергают лизису при охлаждении льдом
буфер. Лизаты клеток обрабатывали ультразвуком и центрифугировали при 12 000 ×
г при 4°С в течение 5 мин. Концентрация белка в экстрактах была
определяется с использованием реагента Бредфорда от Bio-Rad Laboratories.
Равные количества белка (20 мкг/дорожка) растворялись с помощью натрия.
электрофорез в полиакриламидном геле с додецилсульфатом (SDS-PAGE) и
переносятся на нитроцеллюлозные мембраны (Millipore, Bedford, MA,
США).Далее мембрану блокировали в 5% обезжиренном сухом молоке в
Трис-буферный физиологический раствор с твином-20 в течение 2 ч при комнатной температуре и
затем инкубировали с различными первичными антителами в течение ночи при 4°С.
Использовались конъюгированные с HRP вторичные антитела (1:20 000).
затем в течение 2 ч при комнатной температуре. Иммунореактивные полосы были
визуализируется усилением хемилюминесценции (Пирс, Рокфорд, Иллинойс,
США) и количественный анализ с использованием программного обеспечения ImageJ (National Institutes
здравоохранения, Бетесда, Мэриленд, США).
Концентрация белка в экстрактах была
определяется с использованием реагента Бредфорда от Bio-Rad Laboratories.
Равные количества белка (20 мкг/дорожка) растворялись с помощью натрия.
электрофорез в полиакриламидном геле с додецилсульфатом (SDS-PAGE) и
переносятся на нитроцеллюлозные мембраны (Millipore, Bedford, MA,
США).Далее мембрану блокировали в 5% обезжиренном сухом молоке в
Трис-буферный физиологический раствор с твином-20 в течение 2 ч при комнатной температуре и
затем инкубировали с различными первичными антителами в течение ночи при 4°С.
Использовались конъюгированные с HRP вторичные антитела (1:20 000).
затем в течение 2 ч при комнатной температуре. Иммунореактивные полосы были
визуализируется усилением хемилюминесценции (Пирс, Рокфорд, Иллинойс,
США) и количественный анализ с использованием программного обеспечения ImageJ (National Institutes
здравоохранения, Бетесда, Мэриленд, США).
МТТ-анализ
Жизнеспособность клеток оценивали с помощью анализа МТТ.Кратко,
клетки высевали в 48-луночный плоскодонный планшет. (3×104 клеток на лунку) и культивировали в течение ночи. Затем
клетки голодали в течение ночи и обрабатывались различными препаратами, как
указано позже. Для эксперимента с курсовой дозой клетки обрабатывали
с концентрациями гармина и инкубировали 48 ч. Для курса времени
эксперимента клетки инкубировали с 10 мкМ гармина в течение 12, 24,
36 и 48 часов. После инкубации к раствору добавляли 20 мкл МТТ (5 мг/мл).
в каждой лунке, и клеткам давали возможность расти в полной среде при
37°С в течение 3 часов.Супернатант удаляли, затем добавляли 500 мкл ДМСО.
добавляли в каждую лунку и перемешивали в течение 10 мин для растворения кристалла.
Затем определяли оптическую плотность с помощью микропланшета.
считыватель спектрофотометра при 570 нм.
(3×104 клеток на лунку) и культивировали в течение ночи. Затем
клетки голодали в течение ночи и обрабатывались различными препаратами, как
указано позже. Для эксперимента с курсовой дозой клетки обрабатывали
с концентрациями гармина и инкубировали 48 ч. Для курса времени
эксперимента клетки инкубировали с 10 мкМ гармина в течение 12, 24,
36 и 48 часов. После инкубации к раствору добавляли 20 мкл МТТ (5 мг/мл).
в каждой лунке, и клеткам давали возможность расти в полной среде при
37°С в течение 3 часов.Супернатант удаляли, затем добавляли 500 мкл ДМСО.
добавляли в каждую лунку и перемешивали в течение 10 мин для растворения кристалла.
Затем определяли оптическую плотность с помощью микропланшета.
считыватель спектрофотометра при 570 нм.
Анализ заживления ран
клетки SKOV-3 высевали в 6-луночный планшет и
культивируют до конфлюэнтного монослоя. Клетки предварительно обрабатывали
гидроксимочевина (5 ммоль/л) для предотвращения пролиферации клеток. Наконечник пипетки
(200 мкл) использовали для царапания раны по средней линии каждой лунки. затем клетки дважды промывали PBS.После 0, 12, 24,
36 ч культивирования в RPMI-1640 с добавлением 10% сыворотки (контроль)
или гарминовых стимуляторов миграцию клеток оценивали по
измерение разницы в площади ран с помощью Leica
Система анализа изображений DM2500 (Leica, Мангейм, Германия).
затем клетки дважды промывали PBS.После 0, 12, 24,
36 ч культивирования в RPMI-1640 с добавлением 10% сыворотки (контроль)
или гарминовых стимуляторов миграцию клеток оценивали по
измерение разницы в площади ран с помощью Leica
Система анализа изображений DM2500 (Leica, Мангейм, Германия).
Анализ образования колоний
клетки SKOV-3 в логарифмической фазе роста были высевали в 6-луночный планшет и культивировали в среде RPMI-1640 с добавлением 10% FBS на ночь. Затем клетки обрабатывали указанными концентрациями гармина еще две недели.Затем супернатанты удаляли, а клетки тщательно трижды промывали фосфатно-солевым буфером (PBS). Затем клетки фиксировали фиксатором, состоящим из метанол-ледяная уксусная кислота (3:1) в течение 10 мин. Наконец, клетки окрашивали в кристаллический фиолетовый в течение 20 мин. Образы колоний были сняты цифровым фотоаппаратом.
Статистический анализ
Данные указаны как среднее значение ± стандартная ошибка среднего по крайней мере из 3
самостоятельные эксперименты. Статистический анализ проводился с использованием
критерий Стьюдента (сравнение двух групп) или однофакторный анализ
дисперсии (ANOVA) с последующим тестом Даннета или Тьюки
(сравнение более двух групп).Значение p<0,05 было
считаются статистически различными. Все статистические анализы были
проводится с использованием Prism версии 6.0 (программное обеспечение GraphPad).
Статистический анализ проводился с использованием
критерий Стьюдента (сравнение двух групп) или однофакторный анализ
дисперсии (ANOVA) с последующим тестом Даннета или Тьюки
(сравнение более двух групп).Значение p<0,05 было
считаются статистически различными. Все статистические анализы были
проводится с использованием Prism версии 6.0 (программное обеспечение GraphPad).
Результаты
Harmine ингибирует базальную клетку уровень пролиферации в клетках SKOV-3
Сначала мы исследовали влияние гармина на
пролиферация клеток SKOV-3. Клетки SKOV-3 инкубировали
с различной концентрацией гармина (0, 1, 2, 5, 10, 20 и 30
мкМ) в течение 48 ч, а затем жизнеспособность клеток измеряли с помощью анализа МТТ.В виде
показано на рис. 1А, гармин заметно
ингибировал рост клеток SKOV-3 дозозависимым образом, как
концентрация менее 2 мкМ малоэффективна, в то время как
концентрации более 5 мкМ постепенно подавляли рост
клеток SKOV-3 (рис. 1А). Мы следующие
изучали влияние гармина в разное время лечения на
Жизнеспособность клеток SKOV-3, клетки инкубировали с 10 мкМ гармина. на 12, 24, 36 или 48 часов. Анализы жизнеспособности клеток показали, что гармин
значительно ингибировал рост клеток SKOV-3, за исключением 12-часовой инкубации,
с самым сильным эффектом через 48 ч (рис.1Б).
на 12, 24, 36 или 48 часов. Анализы жизнеспособности клеток показали, что гармин
значительно ингибировал рост клеток SKOV-3, за исключением 12-часовой инкубации,
с самым сильным эффектом через 48 ч (рис.1Б).
Для дальнейшего подтверждения ингибирующего действия клеток пролиферации гармина, были проведены анализы образования колоний. Как показано на рис. 1C, последовательный с анализом МТТ гармин значительно подавлял рост колонии образование клеток SKOV-3 дозозависимым образом (рис. 1C и D). Для предоставления дополнительных доказательств, затем мы обнаружили экспрессию пролиферирующих клеток ядерный антиген (PCNA), белковый маркер, который широко используется для индикации пролиферации клеток. Мы обнаружили, что выражение PCNA была значительно снижена при обработке гармином как для 24 или 48 ч (рис.1Е и F). Взятый вместе, наши приведенные выше результаты убедительно показали, что гармин показал значительный ингибирующий эффект на рост клеток в SKOV-3 клетки.
Harmine ингибирует EGF-индуцированный пролиферация клеток SKOV-3
Широко известно, что эпидермальный фактор роста
Рецептор (EGFR) часто гиперэкспрессируется при многих раковых заболеваниях человека. включая рак яичников, и известно, что активация EGFR
участие в росте и прогрессировании различных злокачественных новообразований
(12,13). Затем мы спросили, был ли гармин
способен ингибировать EGF-индуцированную пролиферацию клеток SKOV-3.К
Для этого клетки обрабатывали гармином в отсутствие или
присутствии EGF (50 нг/мл) в течение 48 ч, а затем жизнеспособность клеток
измеряется методом МТТ. Как показано на рис.
2А, EGF значительно увеличивал пролиферацию SKOV-3.
клетки, в то время как эта усиленная EGF пролиферация была заметно
подавляется обработкой гармином (рис.
2А). В соответствии с предыдущими результатами, гармин не только
ингибирует усиленную EGF пролиферацию, но также ингибирует базальную
уровень пролиферации клеток SKOV-3 в отсутствие EGF
(Инжир.2А). Подобные результаты были
наблюдается с помощью анализа образования колоний. Как и ожидалось, гармин
значительно ингибирует EGF-индуцированную способность к образованию колоний
(рис. 2В и С). Эти результаты
еще раз подтвердили, что гармин, очевидно, может ингибировать работу яичников.
включая рак яичников, и известно, что активация EGFR
участие в росте и прогрессировании различных злокачественных новообразований
(12,13). Затем мы спросили, был ли гармин
способен ингибировать EGF-индуцированную пролиферацию клеток SKOV-3.К
Для этого клетки обрабатывали гармином в отсутствие или
присутствии EGF (50 нг/мл) в течение 48 ч, а затем жизнеспособность клеток
измеряется методом МТТ. Как показано на рис.
2А, EGF значительно увеличивал пролиферацию SKOV-3.
клетки, в то время как эта усиленная EGF пролиферация была заметно
подавляется обработкой гармином (рис.
2А). В соответствии с предыдущими результатами, гармин не только
ингибирует усиленную EGF пролиферацию, но также ингибирует базальную
уровень пролиферации клеток SKOV-3 в отсутствие EGF
(Инжир.2А). Подобные результаты были
наблюдается с помощью анализа образования колоний. Как и ожидалось, гармин
значительно ингибирует EGF-индуцированную способность к образованию колоний
(рис. 2В и С). Эти результаты
еще раз подтвердили, что гармин, очевидно, может ингибировать работу яичников. пролиферация раковых клеток.
пролиферация раковых клеток.
Harmine значительно подавляет миграция клеток SKOV-3
Миграция клеток играет важную роль при раке прогресс. Изучить влияние гармина на миграцию Клетки SKOV-3, был проведен типичный анализ заживления царапины. для измерения коэффициента миграции.Как показали наши вышеприведенные результаты, гармин был способен ингибировать пролиферацию клеток SKOV-3, это может помешать анализу результатов миграции клеток. Таким образом, чтобы исключить угнетающее пролиферацию действие гармина на миграции, исследование заживления раны проводили в присутствии гидроксимочевины, которая предотвращает пролиферацию клеток, ингибируя Синтез ДНК. Как показано на рис. 3, гармин значительно ингибировал миграцию клеток SKOV-3 через 24 и через 36 ч после обработки, в то время как через 12 ч эффект был незначительным (рис.3А и В).
Блоки Harmine ERK1/2 и Фосфорилирование CREB в клетках SKOV-3
Мы продемонстрировали, что гармин значительно
подавлял рост клеток SKOV-3. Это подняло вопрос о том, как гармин
ингибирует рост клеток SKOV-3. Митоген-активируемый белок
пути киназы (MAPK) составляют большую модульную сеть, которая
регулирует различные физиологические процессы, в том числе клеточные
рост и дифференциация. Таким образом, мы сначала отслеживали
Уровни фосфорилирования ERK1/2 после гармина
лечение.Как показано на рис. 4,
гармин вызывал быстрое снижение уровня фосфорилирования
ERK1/2 без изменений в сумме ERK1/2
уровни экспрессии (фиг. 4А).
Точно так же уровень фосфорилирования CREB, важный
фактор транскрипции, играющий важную роль в клетке
пролиферация также уменьшилась таким же образом, как и
ERK1/2 (рис. 4Б), который
можно предположить, что гармин ингибирует ERK1/2
фосфорилирование, которое, в свою очередь, подавляло активацию CREB.
Это подняло вопрос о том, как гармин
ингибирует рост клеток SKOV-3. Митоген-активируемый белок
пути киназы (MAPK) составляют большую модульную сеть, которая
регулирует различные физиологические процессы, в том числе клеточные
рост и дифференциация. Таким образом, мы сначала отслеживали
Уровни фосфорилирования ERK1/2 после гармина
лечение.Как показано на рис. 4,
гармин вызывал быстрое снижение уровня фосфорилирования
ERK1/2 без изменений в сумме ERK1/2
уровни экспрессии (фиг. 4А).
Точно так же уровень фосфорилирования CREB, важный
фактор транскрипции, играющий важную роль в клетке
пролиферация также уменьшилась таким же образом, как и
ERK1/2 (рис. 4Б), который
можно предположить, что гармин ингибирует ERK1/2
фосфорилирование, которое, в свою очередь, подавляло активацию CREB.
Harmine ингибирует EGF-индуцированный Фосфорилирование ERK1/2 и CREB в клетках SKOV-3
Гармин значительно ингибировал клетки
пролиферации и фосфорилирования ERK1/2 и CREB в
Клетки SKOV-3, однако, мы не знаем, оказывает ли ингибирующее действие
гармина на пролиферацию клеток опосредовано
Путь ERK1/2/CREB. Чтобы убедиться в этом, мы далее
исследовали, может ли гармин блокировать EGF-индуцированный
Фосфорилирование ERK1/2 и CREB. Как показано на рис. 5, мы сначала подтвердили, что EGF
лечение значительно увеличило фосфорилирование
ERK1/2 и CREB (рис. 5А и
Б). Однако как EGF-индуцированные ERK1/2, так и CREB
фосфорилирование были заметно ингибированы обработкой гармином
(Рис. 5А и В). Показывать
что фосфорилирование CREB было ниже ERK1/2, мы
оценили действие селективного ингибитора EKR1/2
на EGF-индуцированное фосфорилирование CREB в клетках SKOV-3.Клетки были
предварительно обрабатывали в течение 30 минут U0126 (10 мкМ), а затем стимулировали
ЭФР. Мы обнаружили, что U0126 блокирует индуцированные EGF ERK1/2 и
CREB (рис. 5C и D)
фосфорилирование, предполагая, что CREB действовал как нижестоящий фактор
ЭРК1/2.
Чтобы убедиться в этом, мы далее
исследовали, может ли гармин блокировать EGF-индуцированный
Фосфорилирование ERK1/2 и CREB. Как показано на рис. 5, мы сначала подтвердили, что EGF
лечение значительно увеличило фосфорилирование
ERK1/2 и CREB (рис. 5А и
Б). Однако как EGF-индуцированные ERK1/2, так и CREB
фосфорилирование были заметно ингибированы обработкой гармином
(Рис. 5А и В). Показывать
что фосфорилирование CREB было ниже ERK1/2, мы
оценили действие селективного ингибитора EKR1/2
на EGF-индуцированное фосфорилирование CREB в клетках SKOV-3.Клетки были
предварительно обрабатывали в течение 30 минут U0126 (10 мкМ), а затем стимулировали
ЭФР. Мы обнаружили, что U0126 блокирует индуцированные EGF ERK1/2 и
CREB (рис. 5C и D)
фосфорилирование, предполагая, что CREB действовал как нижестоящий фактор
ЭРК1/2.
Harmine уменьшает выраженность VEGF, MMP-2 и MMP-9 в клетках SKOV-3
Поскольку фактор роста эндотелия сосудов (VEGF) и
матриксные металлопротеиназы (ММП), особенно желатиназы
Было показано, что MMP-2 и MMP-9 играют важную роль в
пролиферация, миграция и метастазирование многих первичных эпителиальных
опухоли (14,15), затем мы исследовали влияние
гармина на экспрессию VEGF, MMP-2 и MMP-9 в
Ячейки СКОВ-3. Как показано на фиг. 6А,
ПЦР-анализ с обратной транскрипцией показал, что ген
транскрипция VEGF, MMP-2 и MMP-9 была
значительное подавление в присутствии гармина (10 мкМ) в течение 48
ч (рис. 6А и В). Последовательно,
количественный анализ ПЦР в реальном времени также показал, что гармин
заметно подавлял уровень мРНК VEGF (фиг. 6C), MMP-2 (фиг. 6D) и MMP-9 (фиг. 6E).
Как показано на фиг. 6А,
ПЦР-анализ с обратной транскрипцией показал, что ген
транскрипция VEGF, MMP-2 и MMP-9 была
значительное подавление в присутствии гармина (10 мкМ) в течение 48
ч (рис. 6А и В). Последовательно,
количественный анализ ПЦР в реальном времени также показал, что гармин
заметно подавлял уровень мРНК VEGF (фиг. 6C), MMP-2 (фиг. 6D) и MMP-9 (фиг. 6E).
Обсуждение
В настоящем исследовании мы исследовали противоопухолевый влияние и потенциальный механизм действия гармина на рак яичников человека клетки.Наши результаты показали, что гармин не только значительно ингибирует пролиферацию и миграцию клеток SKOV-3, но также подавлял EGF-индуцированную пролиферацию клеток SKOV-3. Кроме того, этот противоопухолевый эффект гармина может заключаться в ингибировании Путь EKR1/2/CREB и подавление экспрессии VEGF и ММП.
Многие природные алкалоиды, такие как камптотецин,
Винкристин и эллиптицин обладают мощным противораковым действием.
агентов в последние годы (16). Эти
алкалоиды убивают опухолевые клетки различными биохимическими механизмами.
действие, такое как ингибирование апоптоза, ингибирование
топоизомераза I и II и ингибирование образования микротрубочек.Однако молекулярные механизмы противоопухолевой активности
β-карболины не полностью определены. Примечательно, что сообщается, что
некоторые β-карболины и родственные им соединения, по-видимому, обладают
цитотоксическое действие на несколько клеточных линий, включая раковые клетки и
гранулярные нейроны мозжечка. Хотя цитотоксические агенты, вызывающие
гибель клеток и, в конечном итоге, уменьшение размера опухоли являются обычными
противораковые препараты и до сих пор наиболее широко используются во всем мире
(17), нецитотоксический,
молекулярно-направленные препараты, ингибирующие рост опухоли без прямого
цитотоксичность вызывает наибольшую озабоченность (18).В нашем исследовании ингибирующий эффект
роста клеток в присутствии гармина может быть результатом двух
возможные причины, от неспецифического цитотоксического действия гармина
которые быстро убили раковые клетки, или что гармин может подавить
пролиферацию клеток SKOV-3 без индукции острого клеточного
повреждать. Чтобы отличить последнее от первого, мы измерили ячейку
жизнеспособность клеток SKOV-3, обработанных 10 мкМ гармина или без него, для
12 ч. Наши результаты показали, что существенной разницы нет.
между жизнеспособностью контрольных клеток и обработанных гармином
клетки.С другой стороны, недавнее исследование показало, что лечение
с гармином повышенная пролиферация культивируемых нейральных предшественников
клеток без повреждения ДНК или гибели клеток (19). Кроме того, было обнаружено, что гармин
способствовать пролиферации клеток, так как воздействие гармина на куриные эмбрионы
заметно повышенное включение BrdU и количество митотических клеток
в спинном мозге (20). Вместе
с нашими результатами, можно предположить, что ингибирование
пролиферация клеток SKOV-3 под действием гармина может быть не связана с его
неспецифический цитотоксический эффект.
Чтобы отличить последнее от первого, мы измерили ячейку
жизнеспособность клеток SKOV-3, обработанных 10 мкМ гармина или без него, для
12 ч. Наши результаты показали, что существенной разницы нет.
между жизнеспособностью контрольных клеток и обработанных гармином
клетки.С другой стороны, недавнее исследование показало, что лечение
с гармином повышенная пролиферация культивируемых нейральных предшественников
клеток без повреждения ДНК или гибели клеток (19). Кроме того, было обнаружено, что гармин
способствовать пролиферации клеток, так как воздействие гармина на куриные эмбрионы
заметно повышенное включение BrdU и количество митотических клеток
в спинном мозге (20). Вместе
с нашими результатами, можно предположить, что ингибирование
пролиферация клеток SKOV-3 под действием гармина может быть не связана с его
неспецифический цитотоксический эффект.
Наши результаты показали, что гармин ингибирует базальную
уровень пролиферации клеток SKOV-3. Примечательно, что также
значительно подавлял EGF-индуцированную пролиферацию клеток SKOV-3,
предполагая идеальный противоопухолевый эффект гармина. Аберрантный
экспрессия и передача сигналов EGFR-опосредованного пути
связаны с различными видами рака человека, включая рак молочной железы, толстой кишки, головы
рак шеи, поджелудочной железы, легких и яичников. Несомненно, ЭГФР
и члены его семьи стали наиболее полезными биомаркерами
и рациональная цель для лечения рака (21).Стоит отметить, что EGFR был
сообщается, что гиперэкспрессируется в 60% случаев рака яичников.
(22), в то время как повышенная экспрессия
а активация EGFR связана с высоким индексом пролиферации клеток,
высокая скорость миграции и стромальной инвазии (23,24).
Это может в некоторой степени объяснить, почему гармин ингибирует базальную
уровень пролиферации клеток SKOV-3. Однако как гармин влияет
Сигнальный путь EGFR нуждается в дальнейшем изучении.
Аберрантный
экспрессия и передача сигналов EGFR-опосредованного пути
связаны с различными видами рака человека, включая рак молочной железы, толстой кишки, головы
рак шеи, поджелудочной железы, легких и яичников. Несомненно, ЭГФР
и члены его семьи стали наиболее полезными биомаркерами
и рациональная цель для лечения рака (21).Стоит отметить, что EGFR был
сообщается, что гиперэкспрессируется в 60% случаев рака яичников.
(22), в то время как повышенная экспрессия
а активация EGFR связана с высоким индексом пролиферации клеток,
высокая скорость миграции и стромальной инвазии (23,24).
Это может в некоторой степени объяснить, почему гармин ингибирует базальную
уровень пролиферации клеток SKOV-3. Однако как гармин влияет
Сигнальный путь EGFR нуждается в дальнейшем изучении.
Пути митоген-активируемой протеинкиназы (MAPK)
такие как ERK1/2, играют важную роль в регуляции
пролиферации и миграции раковых клеток.В соответствии с
результаты показывают, что гармин ингибирует пролиферацию SKOV-3
клеток, фосфорилирование ERK1/2 было глубоко
блокируется обработкой гармином. Более того, гармин также блокировал
фосфорилирование CREB, которое, как сообщается, подавляет апоптоз,
индуцируют пролиферацию клеток, миграцию и опосредуют метастазирование опухоли
(25). Поскольку способ торможения
фосфорилирования CREB вполне соответствовал
ERK1/2, предполагая, что CREB действует ниже по течению от
ЭРК1/2. Эта гипотеза была подтверждена следующим
данные.Во-первых, мы заметили, что фосфорилирование
ERK1/2 и CREB, опосредованные EGF, были заметно ингибированы.
в присутствии гармина. Во-вторых, EGF-индуцированный CREB
фосфорилирование может быть значительно заблокировано U0126, ингибитором
широко используется для блокирования активности ERK1/2, что указывает на
EGF-индуцированное фосфорилирование CREB опосредуется ERK1/2.
Таким образом, мы заключаем, что ингибирование пути MEK/ERK, который, в свою очередь,
заблокированная активность CREB участвует в подавляющем эффекте гармина
на EGF-индуцированную пролиферацию клеток при раке яичников человека
клетки.
Более того, гармин также блокировал
фосфорилирование CREB, которое, как сообщается, подавляет апоптоз,
индуцируют пролиферацию клеток, миграцию и опосредуют метастазирование опухоли
(25). Поскольку способ торможения
фосфорилирования CREB вполне соответствовал
ERK1/2, предполагая, что CREB действует ниже по течению от
ЭРК1/2. Эта гипотеза была подтверждена следующим
данные.Во-первых, мы заметили, что фосфорилирование
ERK1/2 и CREB, опосредованные EGF, были заметно ингибированы.
в присутствии гармина. Во-вторых, EGF-индуцированный CREB
фосфорилирование может быть значительно заблокировано U0126, ингибитором
широко используется для блокирования активности ERK1/2, что указывает на
EGF-индуцированное фосфорилирование CREB опосредуется ERK1/2.
Таким образом, мы заключаем, что ингибирование пути MEK/ERK, который, в свою очередь,
заблокированная активность CREB участвует в подавляющем эффекте гармина
на EGF-индуцированную пролиферацию клеток при раке яичников человека
клетки.
Деградация внеклеточного матрикса (ECM) и
базальные мембраны опухолевыми клетками является важным шагом в
процессы опухолевой инвазии и метастазирования (26). Матриксные металлопротеиназы (ММП)
семейство протеолитических ферментов, регулирующих различные клетки
поведение, имеющее отношение к биологии рака, посредством деградации
внеклеточный матрикс, окружающий опухоли (14). В нашем исследовании мы обнаружили, что гармин
может ингибировать экспрессию MMP-2 и MMP-9. Рано
Адгезия и метастазирование клеток рака яичников опосредованы
Семейство белков MMP, экспрессия которых усилена
во время прогрессирования рака яичников (27).Хотя MMP считались
тесно коррелировать с инвазией опухоли и метастазированием (27), исследования также показывают, что ингибирование
действия ММП-2 и ММП-9 могут привести к
снижение ремоделирования ВКМ и подавление эндотелиальных клеток
пролиферация и миграция (28).
Более того, в соответствии с нашими результатами, недавнее исследование указывает
что активация пути ERK1/2/CREB способствует
экспрессия MMP-2 и MMP-9, что может объяснить
пролиферация клеток рака легкого (29). С другой стороны, наши результаты также
показали, что гармин может ингибировать экспрессию VEGF.
Матриксные металлопротеиназы (ММП)
семейство протеолитических ферментов, регулирующих различные клетки
поведение, имеющее отношение к биологии рака, посредством деградации
внеклеточный матрикс, окружающий опухоли (14). В нашем исследовании мы обнаружили, что гармин
может ингибировать экспрессию MMP-2 и MMP-9. Рано
Адгезия и метастазирование клеток рака яичников опосредованы
Семейство белков MMP, экспрессия которых усилена
во время прогрессирования рака яичников (27).Хотя MMP считались
тесно коррелировать с инвазией опухоли и метастазированием (27), исследования также показывают, что ингибирование
действия ММП-2 и ММП-9 могут привести к
снижение ремоделирования ВКМ и подавление эндотелиальных клеток
пролиферация и миграция (28).
Более того, в соответствии с нашими результатами, недавнее исследование указывает
что активация пути ERK1/2/CREB способствует
экспрессия MMP-2 и MMP-9, что может объяснить
пролиферация клеток рака легкого (29). С другой стороны, наши результаты также
показали, что гармин может ингибировать экспрессию VEGF. Поскольку VEGF является наиболее важным фактором митогенеза и выживания
участвует в ангиогенезе, в то время как ангиогенез необходим для
рост солидных опухолей, нацеливание на сосуды было исследовано как
потенциальная стратегия для подавления роста опухоли и метастазирования. Таким образом,
этот результат еще раз подтвердил противоопухолевый эффект гармина.
Однако, способен ли гармин ингибировать ангиогенез в
развитие солидной опухоли требует дальнейшего изучения.
Поскольку VEGF является наиболее важным фактором митогенеза и выживания
участвует в ангиогенезе, в то время как ангиогенез необходим для
рост солидных опухолей, нацеливание на сосуды было исследовано как
потенциальная стратегия для подавления роста опухоли и метастазирования. Таким образом,
этот результат еще раз подтвердил противоопухолевый эффект гармина.
Однако, способен ли гармин ингибировать ангиогенез в
развитие солидной опухоли требует дальнейшего изучения.
Благодарности
Настоящее исследование было поддержано грантами Исследования традиционной китайской медицины (номер.2016B032), Наука и Проект технологического планирования (№ 20175032) провинции Цзянси Комиссия по здравоохранению и планированию семьи, Руководство по науке и Технологическая программа Наньчана (№ 2016-ZDXXM-039).
Глоссарий
Сокращения
Сокращения:
КРЭБ | циклический аденозинмонофосфат белок, связывающий элемент ответа |
ЕСМ | внеклеточный матрикс |
ЭФР | эпидермальный фактор роста |
ЭГФР | Рецептор эпителиального фактора роста |
ЭРК 1/2 | киназа, регулируемая внеклеточным сигналом 1/2 |
МАПК | митоген-активируемая протеинкиназа |
ММП | матриксная металлопротеиназа |
ВЭФР | фактор роста эндотелия сосудов |
Ссылки
 w3.org/1999/xhtml»>
w3.org/1999/xhtml»>1
Торре Л.А., Брей Ф., Сигел Р.Л., Ферлей Дж., Лорте-Тиулент Дж. и Джемаль А.: Глобальная статистика рака, 2012 г.Калифорния Рак Дж. Клин. 65:87–108. 2015. Просмотр статьи: Google Scholar: PubMed/NCBI
.2
Коулман Р.Л., Монк Б.Дж., Суд А.К. и Херцог TJ: Последние исследования и лечение эпителия на поздних стадиях рак яичников. Nat Rev Clin Oncol. 10: 211–224. 2013. Просмотр статьи: Google Scholar: PubMed/NCBI
.3
Мукерджи А.К., Басу С., Саркар Н. и Гош AC: Достижения в лечении рака с помощью натуральных продуктов на растительной основе.Курр Мед Хим. 8: 1467–1486. 2001. Просмотр статьи: Google Scholar: PubMed/NCBI
.4
Макчесни Д. Д.: Натуральные продукты в лекарствах
открытие — организация успеха. PR Health Sci J. 21:91–95.
2002. PubMed/NCBI
Д.: Натуральные продукты в лекарствах
открытие — организация успеха. PR Health Sci J. 21:91–95.
2002. PubMed/NCBI
5
Cragg GM, Newman DJ и Snader KM: Natural продукты в области открытия и разработки лекарств. J Nat Prod. 60:52–60. 1997. Просмотр статьи: Google Scholar: PubMed/NCBI
.6
Цао Р, Пэн В, Ван Цзи и Сюй А: бета-карболиновые алкалоиды: биохимические и фармакологические функции.Курр Мед Хим. 14: 479–500. 2007. Просмотр статьи: Google Scholar: PubMed/NCBI
.7
Патель К., Гадевар М., Трипати Р., Прасад С.К.
и Патель Д.К.: Обзор медицинского значения, фармакологических
активность и биоаналитические аспекты бета-карболинового алкалоида
«Гармине». Asian Pac J Trop Biomed. 2: 660–664.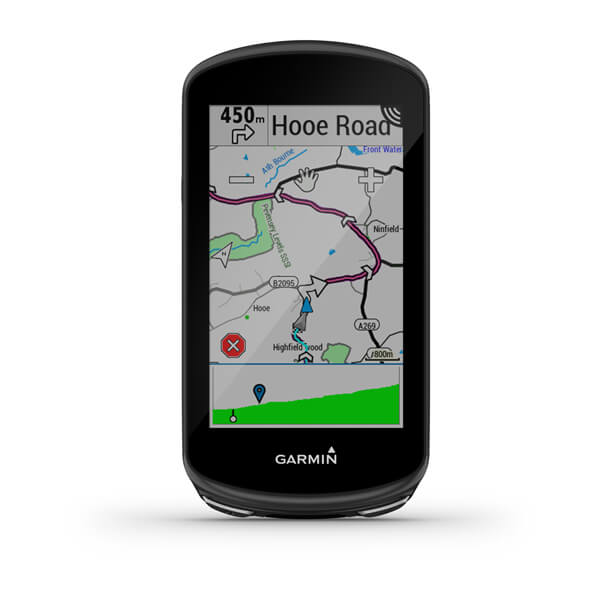 2012. Просмотр статьи: Google Scholar: PubMed/NCBI
2012. Просмотр статьи: Google Scholar: PubMed/NCBI
8
Фарзин Д. и Мансури Н.: Антидепрессантоподобный эффект гармана и других бета-карболинов у тест принудительного плавания мыши.Евро Нейропсихофармакол. 16:324–328. 2006. Просмотр статьи: Google Scholar: PubMed/NCBI
.9
Цао М.Р., Ли К., Лю З.Л., Лю Х.Х., Ван В., Ляо XL, Pan YL и Jiang JW: Harmine индуцирует апоптоз в клетках HepG2 через митохондриальный сигнальный путь. Гепатобилиарная недостаточность поджелудочной железы Междунар. 10: 599–604. 2011. Просмотр статьи: Google Scholar: PubMed/NCBI
.10
Hamsa TP и Kuttan G: Harmine активируется внутренние и внешние пути апоптоза при меланоме B16F-10.Чин Мед. 6:112011. Просмотр статьи : Академия Google : PubMed/NCBI
11
Чжан П. , Хуан Ч.Р., Ван В., Чжан С.К., Чен
JJ, Wang JJ, Lin C и Jiang JW: Гидрохлорид гармина вызывает G2
фаза остановки и апоптоза в клетках MGC-803 и клетках SMMC-7721 путем
повышающая регуляция p21, активирующая каспазу-8/Bid и понижающая регуляция
ERK/Плохой путь. Фитотер Рез. 30:31–40. 2016. Просмотр статьи: Google Scholar: PubMed/NCBI
, Хуан Ч.Р., Ван В., Чжан С.К., Чен
JJ, Wang JJ, Lin C и Jiang JW: Гидрохлорид гармина вызывает G2
фаза остановки и апоптоза в клетках MGC-803 и клетках SMMC-7721 путем
повышающая регуляция p21, активирующая каспазу-8/Bid и понижающая регуляция
ERK/Плохой путь. Фитотер Рез. 30:31–40. 2016. Просмотр статьи: Google Scholar: PubMed/NCBI
12
Лафки Дж. М., Уилкен Дж. А., Барон А. Т. и Майле Нью-Джерси: Клинические последствия ErbB/эпидермального фактора роста (EGF) Семейство рецепторов и его лиганды при раке яичников.Биохим Биофиз Акта. 1785: 232–265. 2008. PubMed/NCBI
.13
Лю LZ, Hu XW, Xia C, He J, Zhou Q, Shi X,
Fang J и Jiang BH: Реактивные формы кислорода регулируют эпидермальный
индуцированный фактором роста фактор роста эндотелия сосудов и
экспрессия индуцируемого гипоксией фактора-1альфа посредством активации
AKT и P70S6K1 в клетках рака яичников человека. Свободный Радик Биол Мед.
41:1521–1533. 2006. Просмотр статьи: Google Scholar: PubMed/NCBI
Свободный Радик Биол Мед.
41:1521–1533. 2006. Просмотр статьи: Google Scholar: PubMed/NCBI
14
Newby AC: матриксные металлопротеиназы регулируют миграцию, пролиферацию и гибель сосудистых гладких мышечные клетки путем деградации матриксных и нематричных субстратов.Кардиовасц Рез. 69:614–624. 2006. Просмотр статьи: Google Scholar: PubMed/NCBI
.15
Берначес П.Н., Сокер С. и Сируа М.Г.: Влияние сосудистого эндотелиального фактора роста на эндотелиальные клетки пролиферация, миграция и синтез фактора активации тромбоцитов является Flk-1-зависимым. Дж. Биол. Хим. 274:31047–31054. 1999. Просмотр статьи: Google Scholar: PubMed/NCBI
.16
Демен А.Л. и Вайшнав П.: Натуральные продукты
для химиотерапии рака. Микроб Биотехнология. 4: 687–699. 2011.
Просмотр статьи : Академия Google : PubMed/NCBI
Микроб Биотехнология. 4: 687–699. 2011.
Просмотр статьи : Академия Google : PubMed/NCBI
17
Куммар С., Гутьеррес М., Дорошоу Дж. Х. и Мурго А.Дж.: Разработка лекарств в онкологии: классические цитотоксические и молекулярно-направленные агенты. Бр Дж Клин Фармакол. 62:15–26. 2006. Просмотр статьи : Академия Google : PubMed/NCBI
18
Fox E, Curt GA и Balis FM: Клинические исследования дизайн исследования для таргетной терапии.Онколог. 7: 401–409. 2002. Просмотр статьи : Академия Google : PubMed/NCBI
19
Дакич В., Масиэль Р.М., Драммонд Х., Насименто JM, Trindade P и Rehen SK: Harmine стимулирует пролиферацию предшественники нейронов человека. Пир Дж. 4:e27272016. Просмотр статьи : Академия Google : PubMed/NCBI
20
Хаммерле Б. , Улин Э., Гимера Дж., Беккер В.,
Guillemot F и Tejdor FJ: переходная экспрессия Mnb/Dyrk1a
соединяет выход из клеточного цикла и дифференцировку предшественников нейронов
вызывая экспрессию p27KIP1 и подавляя передачу сигналов NOTCH.Разработка. 138: 2543–2554. 2011. Просмотр статьи: Google Scholar: PubMed/NCBI
, Улин Э., Гимера Дж., Беккер В.,
Guillemot F и Tejdor FJ: переходная экспрессия Mnb/Dyrk1a
соединяет выход из клеточного цикла и дифференцировку предшественников нейронов
вызывая экспрессию p27KIP1 и подавляя передачу сигналов NOTCH.Разработка. 138: 2543–2554. 2011. Просмотр статьи: Google Scholar: PubMed/NCBI
21
Сешачарьюлу П., Поннусамы М.П., Харидас Д., Джайн М., Ганти А.К. и Батра С.К.: Нацеливание на сигнальный путь EGFR в терапии рака. Экспертное мнение по этим целям. 16:15–31. 2012. Просмотр статьи : Академия Google : PubMed/NCBI
22
Димова И, Захариева Б, Райчева С, Димитров Р., Доганов Н. и Тончева Д.: Тканевой микрочиповый анализ Изменения числа копий EGFR и erbB2 в опухолях яичников.Инт Дж. Гинекол Рак. 16:145–151. 2006. Просмотр статьи: Google Scholar: PubMed/NCBI
.
23
Дэнси Дж. Э. и Фрейдлин Б.: Таргетинг Рецептор эпидермального фактора роста — мы промахнулись? Ланцет. 362:62–64. 2003. Просмотр статьи: Google Scholar: PubMed/NCBI
.24
Лассус Х., Сихто Х., Леминен А., Йоэнсуу Х., Изола Дж., Нуппонен Н.Н. и Буцов Р.: Амплификация генов, мутация, экспрессия белка EGFR и мутации ERBB2 в серозных рак яичников.J Mol Med (Берл). 84:671–681. 2006. Просмотр статьи: Google Scholar: PubMed/NCBI
.25
Aggarwal S, Kim SW, Ryu SH, Chung WC и Koo JS: Подавление роста клеток рака легких путем воздействия на циклические Белок, связывающий элемент ответа AMP. Рак рез. 68:981–988. 2008. Просмотр статьи : Академия Google : PubMed/NCBI
26
Лу П. , Уивер В.М. и Верб З.:
внеклеточный матрикс: динамическая ниша в развитии рака.J-ячейка
биол. 196: 395–406. 2012. Просмотр статьи: Google Scholar: PubMed/NCBI
, Уивер В.М. и Верб З.:
внеклеточный матрикс: динамическая ниша в развитии рака.J-ячейка
биол. 196: 395–406. 2012. Просмотр статьи: Google Scholar: PubMed/NCBI
27
Egeblad M и Werb Z: новые функции для матриксные металлопротеиназы при прогрессировании рака. Нат Рев Рак. 2: 161–174. 2002. Посмотреть Статья : Google Scholar : PubMed/NCBI
28
Гомес Д.Э., Алонсо Д.Ф., Йошиджи Х. и Thorgeirsson UP: Тканевые ингибиторы металлопротеиназ: Структура, регуляция и биологические функции.Eur J Cell Biol. 74:111–122. 1997. PubMed/NCBI
.29
Ху П., Хе Дж., Лю С., Ван М., Пань Б. и Чжан
W: Активация β2-адренорецепторов способствует пролиферации
Клетки рака легкого A549 через путь ERK1/2/CREB. Онкол Респ. 36: 1757–1763. 2016. Просмотр статьи: Google Scholar: PubMed/NCBI
36: 1757–1763. 2016. Просмотр статьи: Google Scholar: PubMed/NCBI
Поставщик Harmine | КАС 343-27-1
Harmine (343-27-1) представляет собой селективный конкурентный и обратимый ингибитор МАО-А (IC 50 = 8 мкМ). 1 Гармин является мощным и специфическим ингибитором как киназной активности, так и аутофосфорилирования тирозина во время трансляции DYRK1A (IC 50 : DYRKA1A = 33 нМ; DYRKA1B = 166 нМ; DYRK2 = 1,9 мкМ; DYRK4 = 80 мкМ) . 2,3 Ингибирует самообновление и способствует дифференцировке стволовых клеток глиобластомы (GSLC), возможно, посредством ингибирования фосфорилирования Akt. 4 Гармин также обладает антимикробными, противогрибковыми, антиплазмодиальными, противоопухолевыми, антиоксидантными, антимутагенными и антигенотоксическими свойствами. 5 Ингибирует дифференцировку остеокластов и резорбцию кости за счет подавления c-Fos и NFATc1, индуцированного RANKL. 6
Ссылки/Цитаты:
1) Wouters (1998) Structural Aspects of Monoamine Oxidase and its Reversible Inhibition Current Medicinal Chemistry, 5 137
(2008) DYRK1A фосфорилирует каспазу 9 в ингибирующем сайте и сильно ингибируется в клетках человека гармином ; ФЕБС Дж. , 275 6268
, 275 6268
3) Гоклер и др. . (2009) Harmine специфически ингибирует протеинкиназу DYRK1A и препятствует росту нейритов ; FEBS J., 276 6324
4) Liu et al . (2013) Хармина гидрохлорид ингибирует фосфорилирование Akt и истощает пул раковых стволовых клеток глиобластомы ; J.Neurooncol., 112 39
5) Patel et al . (2012) Обзор медицинского значения, фармакологической активности и биоаналитических аспектов бета-карболинового алкалоида «Гармин» ; Азиатский пакет.Дж. Троп. Biomed., 2 660
6) Yoneazwa et al . (2011) Гармин, β-карболиновый алкалоид, ингибирует дифференцировку остеокластов и резорбцию костей in vitro и in vivo ; Евро. J. Pharmacol., 650 511
Алкалоиды гармала обыкновенного: противораковые биомаркеры с промизином…: Ingenta Connect
Справочная информация: Рак является серьезной и растущей глобальной проблемой здравоохранения во всем мире. В раковых клетках нормальный клеточный цикл был нарушен серией необратимых изменений.Недавние исследования в области фитотерапии и выяснения потенциала фитохимических веществ в лечении рака
был увеличен. Сочетание фитохимических веществ с традиционными подходами к лечению рака может улучшить результаты за счет ускорения гибели клеток, сдерживания пролиферации и инвазии клеток, повышения чувствительности раковых клеток и укрепления иммунной системы. Таким образом, фитохимические вещества могут быть введены
в качестве соответствующих дополнительных лекарственных средств в терапии рака. Peganum Harmala L. (Zygophyllaceae) как ценное лекарственное растение содержит различные алкалоидные ингредиенты.
В раковых клетках нормальный клеточный цикл был нарушен серией необратимых изменений.Недавние исследования в области фитотерапии и выяснения потенциала фитохимических веществ в лечении рака
был увеличен. Сочетание фитохимических веществ с традиционными подходами к лечению рака может улучшить результаты за счет ускорения гибели клеток, сдерживания пролиферации и инвазии клеток, повышения чувствительности раковых клеток и укрепления иммунной системы. Таким образом, фитохимические вещества могут быть введены
в качестве соответствующих дополнительных лекарственных средств в терапии рака. Peganum Harmala L. (Zygophyllaceae) как ценное лекарственное растение содержит различные алкалоидные ингредиенты.
Цель: указание на важность новых способов лечения рака и P.harmala убедительного эффекта в этом
В этом обзоре была предпринята попытка собрать основу для описания возможных масштабов разработки новых лекарств для лечения рака.
Методы: Поиск по ключевым словам «Peganumharmala» и рак, или химиотерапия, или противоопухолевый по базе «Scopus» до 29 th от февраля 2020 г. Были исключены статьи, связанные с сельским хозяйством, химией, науками об окружающей среде и генетике, и были выбраны статьи, посвященные раку.
Были исключены статьи, связанные с сельским хозяйством, химией, науками об окружающей среде и генетике, и были выбраны статьи, посвященные раку.
Результаты и обсуждение: в текущем исследовании 42 статьи, связанные с лечением рака, и 22 статьи о биоактивных компонентах алкалоидов.
собраны из 72 статей. Алкалоиды β-карболина, полученные из P.harmala , особенно гармин, демонстрируют заметные противораковые свойства, воздействуя на апоптоз, аутофагию, аномальную пролиферацию клеток, ангиогенез, метастазирование и цитотоксичность. На основании собранной информации, P.harmala обладает значительной противораковой активностью.Учитывая механизм действия различных противоопухолевых препаратов и их действующее сходство с P.harmala , алкалоиды, полученные из этого растения, в частности гармин, могут быть введены в качестве нового противоракового лекарства отдельно или в качестве адъюванта.
терапия рака.
Нет ссылок
Нет цитирований
Без дополнительных данных
Нет статьи Носитель
Нет показателей
Ключевые слова: Противоопухолевый; Пеганум гармала; рак; химиотерапия; гармин; фитохимический
Тип документа: Обзор статьи
Дата публикации:
1 января 2021 г.
Эта статья была размещена в Интернете 15 декабря 2020 г. в виде статьи Fast Track под названием «Алкалоиды гармалы пегановой: противораковые биомаркеры с многообещающими результатами».
Подробнее об этой публикации?- Current Pharmaceutical Design публикует своевременные подробные обзоры, охватывающие все аспекты текущих исследований в области рационального дизайна лекарств. Каждый выпуск посвящен одному крупному терапевтическому направлению. Приглашенный редактор, который является признанным авторитетом в области терапии, запрашивает для каждого выпуска всесторонние и своевременные обзоры от ведущих исследователей фармацевтической промышленности и научных кругов.
Каждый тематический выпуск Current Pharmaceutical Design охватывает все предметные области, имеющие большое значение для современного дизайна лекарств, включая медицинскую химию, фармакологию, мишени для лекарств и механизм заболевания.
- Редакция
- Информация для авторов
- Подписаться на этот заголовок
- Ingenta Connect не несет ответственности за содержание или доступность внешних веб-сайтов
Первый в своем классе ингибитор скручивания 1 с активностью в отношении рака легких, вызванного онкогенами — Университет Джонса Хопкинса
TY-JOUR
T1 — Первый в своем классе ингибитор скручивания 1 с активностью в отношении рака легких, вызванного онкогенами
AU — Йохум, Захари А.
AU — Cades, Jessica
AU — Mazzacurati, Lucia
AU — Neumann, Neil M.
Au — Kheetarpal, Sushel K.
AU — Chatterjee, Suman
AU — Ван, Hailun
AU — Аттар, Мириам А.
AU — Хуанг, Эрик Х.Б.
AU — Чатли, Сара Н.
AU — Наджент, Катриана
AU — Сомасундарам, Эшвин
AU — Энг, Джонатан А.
AU — Эвальд, Эндрю Дж.
AU
AU — Рудин, Чарльз М.
AU — Тран, Фуок Т.
AU — Бернс, Тимоти Ф.
N1 — Информация о финансировании:
СМ. Рудин является консультантом/членом консультативного совета компаний Abbvie, AstraZeneca, BMS, G1 Therapeutics и Harpoon Therapeutics. П.Т. Тран сообщает о получении коммерческого исследовательского гранта от Medivation Inc-Astellas Pharma, Inc. Другие авторы не сообщили о потенциальных конфликтах интересов. Информация о финансировании:
Работа выполнена при поддержке следующих источников финансирования: Т.Ф. Бернс получил финансирование исследований для этого проекта от премии Американской ассоциации легких (LCD 257864), премии V Foundation Scholar, Фонда Сидни Киммела (SKF-15-099), премии клинического ученого Благотворительного фонда Дорис Дьюк (2015097) и UPCI LUNG. СПОРА CDA P50CA0
СПОРА CDA P50CA0
PY — 2017/12
Y1 — 2017/12
N2 — TWIST1, транскрипционный фактор эпителиально-мезенхимального перехода (EMT), имеет решающее значение для онкоген-управляемого немелкоклеточного рака легкого (NSCLC). Учитывая потенциал TWIST1 в качестве терапевтической мишени, для идентификации ингибиторов TWIST1 был использован химико-биоинформационный подход с использованием анализа картирования связности (CMAP). Характеристика лучших кандидатов из объективного скрининга показала, что гармин, алкалоид гармалы, ингибирует несколько функций TWIST1, включая диссеминацию одиночных клеток, подавление нормального ветвления в 3D эпителиальной культуре и пролиферацию клеток НМРЛ, определяемых драйвером онкогена. Лечение гармином фенокопировало генетическую потерю TWIST1, индуцируя индуцированное онкогеном старение или апоптоз.Механистическое исследование показало, что гармин нацелен на путь TWIST1, способствуя деградации белка TWIST1. Поскольку димеризация имеет решающее значение для функции и стабильности TWIST1, было изучено влияние гармина на специфические димеры TWIST1. TWIST1 и его димерные партнеры, белки E2A, которые, как было установлено, необходимы для TWIST1-опосредованных функций, посттрансляционно регулируют стабильность другого гетеродимерного партнера.
Учитывая потенциал TWIST1 в качестве терапевтической мишени, для идентификации ингибиторов TWIST1 был использован химико-биоинформационный подход с использованием анализа картирования связности (CMAP). Характеристика лучших кандидатов из объективного скрининга показала, что гармин, алкалоид гармалы, ингибирует несколько функций TWIST1, включая диссеминацию одиночных клеток, подавление нормального ветвления в 3D эпителиальной культуре и пролиферацию клеток НМРЛ, определяемых драйвером онкогена. Лечение гармином фенокопировало генетическую потерю TWIST1, индуцируя индуцированное онкогеном старение или апоптоз.Механистическое исследование показало, что гармин нацелен на путь TWIST1, способствуя деградации белка TWIST1. Поскольку димеризация имеет решающее значение для функции и стабильности TWIST1, было изучено влияние гармина на специфические димеры TWIST1. TWIST1 и его димерные партнеры, белки E2A, которые, как было установлено, необходимы для TWIST1-опосредованных функций, посттрансляционно регулируют стабильность другого гетеродимерного партнера. Гармин преимущественно способствовал деградации гетеродимера TWIST1-E2A по сравнению с гомодимером TWIST-TWIST1, и для цитотоксичности гармина требовалось нацеливание на гетеродимер TWIST1-E2A.Наконец, гармин обладал активностью как в трансгенных, так и в ксенотрансплантатных мышиных моделях KRAS-мутантного НМРЛ. Эти исследования идентифицировали гармин как первый в своем классе ингибитор TWIST1 с выраженной противоопухолевой активностью при НМРЛ, вызванном онкогенами, включая мутантный EGFR, мутантный KRAS и НМРЛ с измененным MET. Последствия: TWIST1 необходим для онкоген-управляемого онкогенеза НМРЛ и ЕМТ; таким образом, гармин и его аналоги/производные представляют собой новую терапевтическую стратегию для лечения онкогенного НМРЛ, а также других солидных злокачественных опухолей.
Гармин преимущественно способствовал деградации гетеродимера TWIST1-E2A по сравнению с гомодимером TWIST-TWIST1, и для цитотоксичности гармина требовалось нацеливание на гетеродимер TWIST1-E2A.Наконец, гармин обладал активностью как в трансгенных, так и в ксенотрансплантатных мышиных моделях KRAS-мутантного НМРЛ. Эти исследования идентифицировали гармин как первый в своем классе ингибитор TWIST1 с выраженной противоопухолевой активностью при НМРЛ, вызванном онкогенами, включая мутантный EGFR, мутантный KRAS и НМРЛ с измененным MET. Последствия: TWIST1 необходим для онкоген-управляемого онкогенеза НМРЛ и ЕМТ; таким образом, гармин и его аналоги/производные представляют собой новую терапевтическую стратегию для лечения онкогенного НМРЛ, а также других солидных злокачественных опухолей.
AB — TWIST1, фактор транскрипции эпителиально-мезенхимального перехода (ЭМП), имеет решающее значение для онкогенного онкогенеза немелкоклеточного рака легкого (НМРЛ). Учитывая потенциал TWIST1 в качестве терапевтической мишени, для идентификации ингибиторов TWIST1 был использован химико-биоинформационный подход с использованием анализа картирования связности (CMAP). Характеристика лучших кандидатов из объективного скрининга показала, что гармин, алкалоид гармалы, ингибирует несколько функций TWIST1, включая диссеминацию одиночных клеток, подавление нормального ветвления в 3D эпителиальной культуре и пролиферацию клеток НМРЛ, определяемых драйвером онкогена.Лечение гармином фенокопировало генетическую потерю TWIST1, индуцируя индуцированное онкогеном старение или апоптоз. Механистическое исследование показало, что гармин нацелен на путь TWIST1, способствуя деградации белка TWIST1. Поскольку димеризация имеет решающее значение для функции и стабильности TWIST1, было изучено влияние гармина на специфические димеры TWIST1. TWIST1 и его димерные партнеры, белки E2A, которые, как было установлено, необходимы для TWIST1-опосредованных функций, посттрансляционно регулируют стабильность другого гетеродимерного партнера.Гармин преимущественно способствовал деградации гетеродимера TWIST1-E2A по сравнению с гомодимером TWIST-TWIST1, и для цитотоксичности гармина требовалось нацеливание на гетеродимер TWIST1-E2A.
Характеристика лучших кандидатов из объективного скрининга показала, что гармин, алкалоид гармалы, ингибирует несколько функций TWIST1, включая диссеминацию одиночных клеток, подавление нормального ветвления в 3D эпителиальной культуре и пролиферацию клеток НМРЛ, определяемых драйвером онкогена.Лечение гармином фенокопировало генетическую потерю TWIST1, индуцируя индуцированное онкогеном старение или апоптоз. Механистическое исследование показало, что гармин нацелен на путь TWIST1, способствуя деградации белка TWIST1. Поскольку димеризация имеет решающее значение для функции и стабильности TWIST1, было изучено влияние гармина на специфические димеры TWIST1. TWIST1 и его димерные партнеры, белки E2A, которые, как было установлено, необходимы для TWIST1-опосредованных функций, посттрансляционно регулируют стабильность другого гетеродимерного партнера.Гармин преимущественно способствовал деградации гетеродимера TWIST1-E2A по сравнению с гомодимером TWIST-TWIST1, и для цитотоксичности гармина требовалось нацеливание на гетеродимер TWIST1-E2A. Наконец, гармин обладал активностью как в трансгенных, так и в ксенотрансплантатных мышиных моделях KRAS-мутантного НМРЛ. Эти исследования идентифицировали гармин как первый в своем классе ингибитор TWIST1 с выраженной противоопухолевой активностью при НМРЛ, вызванном онкогенами, включая мутантный EGFR, мутантный KRAS и НМРЛ с измененным MET. Последствия: TWIST1 необходим для онкоген-управляемого онкогенеза НМРЛ и ЕМТ; таким образом, гармин и его аналоги/производные представляют собой новую терапевтическую стратегию для лечения онкогенного НМРЛ, а также других солидных злокачественных опухолей.
Наконец, гармин обладал активностью как в трансгенных, так и в ксенотрансплантатных мышиных моделях KRAS-мутантного НМРЛ. Эти исследования идентифицировали гармин как первый в своем классе ингибитор TWIST1 с выраженной противоопухолевой активностью при НМРЛ, вызванном онкогенами, включая мутантный EGFR, мутантный KRAS и НМРЛ с измененным MET. Последствия: TWIST1 необходим для онкоген-управляемого онкогенеза НМРЛ и ЕМТ; таким образом, гармин и его аналоги/производные представляют собой новую терапевтическую стратегию для лечения онкогенного НМРЛ, а также других солидных злокачественных опухолей.
UR — http://www.scopus.com/inward/record.url?scp=85039870015&partnerID=8YFLogxK
UR — http://www.scopus.com/inward/citedby.url?scp=85039870015&partnerID=8YFLogxK
U2 — 10.1158 / 1541-7786.mcr-17-0298
do — 10.1158 / 1541-7786.mcr-17-0298
м3 — Статья
C2 — 28851812
AN — SCOPUS: 85039870015
VL — 15
SP — 1764
EP — 1776
EP — 1776
Jo — рост клеток и дифференцировка
JF — рост клеток и дифференцирование
SN — 1541-7786
IS — 12
ER —
% PDF -1. 4
%
695 0 объект
>
эндообъект
внешняя ссылка
695 97
0000000016 00000 н
0000002291 00000 н
0000002413 00000 н
0000002919 00000 н
0000003133 00000 н
0000003629 00000 н
0000003757 00000 н
0000004861 00000 н
0000005130 00000 н
0000005151 00000 н
0000005275 00000 н
0000005296 00000 н
0000005412 00000 н
0000005434 00000 н
0000005779 00000 н
0000005802 00000 н
0000007538 00000 н
0000007561 00000 н
0000009335 00000 н
0000009358 00000 н
0000011123 00000 н
0000011407 00000 н
0000011676 00000 н
0000011699 00000 н
0000013568 00000 н
0000013589 00000 н
0000013610 00000 н
0000013633 00000 н
0000015357 00000 н
0000015380 00000 н
0000016985 00000 н
0000017008 00000 н
0000018623 00000 н
0000018646 00000 н
0000020289 00000 н
0000020310 00000 н
0000020402 00000 н
0000020423 00000 н
0000020515 00000 н
0000020537 00000 н
0000021600 00000 н
0000021622 00000 н
0000022152 00000 н
0000022174 00000 н
0000023284 00000 н
0000023307 00000 н
0000025921 00000 н
0000025943 00000 н
0000026829 00000 н
0000026852 00000 н
0000030807 00000 н
0000030830 00000 н
0000033560 00000 н
0000033583 00000 н
0000037208 00000 н
0000037231 00000 н
0000042821 00000 н
0000042844 00000 н
0000047500 00000 н
0000047523 00000 н
0000052914 00000 н
0000052937 00000 н
0000058591 00000 н
0000058614 00000 н
0000063923 00000 н
0000063946 00000 н
0000068651 00000 н
0000068674 00000 н
0000074485 00000 н
0000074508 00000 н
0000080156 00000 н
0000080179 00000 н
0000084762 00000 н
0000084785 00000 н
0000089916 00000 н
0000089939 00000 н
0000094739 00000 н
0000094762 00000 н
0000099794 00000 н
0000099817 00000 н
0000104530 00000 н
0000104553 00000 н
0000110328 00000 н
0000110351 00000 н
0000114046 00000 н
0000114069 00000 н
0000119022 00000 н
0000119045 00000 н
0000123331 00000 н
0000123354 00000 н
0000129428 00000 н
0000129451 00000 н
0000134783 00000 н
0000134806 00000 н
0000140299 00000 н
0000002477 00000 н
0000002897 00000 н
трейлер
]
>>
startxref
0
%%EOF
696 0 объект
>
эндообъект
697 0 объект
>
эндообъект
790 0 объект
>
ручей
Hb««\ À
4
%
695 0 объект
>
эндообъект
внешняя ссылка
695 97
0000000016 00000 н
0000002291 00000 н
0000002413 00000 н
0000002919 00000 н
0000003133 00000 н
0000003629 00000 н
0000003757 00000 н
0000004861 00000 н
0000005130 00000 н
0000005151 00000 н
0000005275 00000 н
0000005296 00000 н
0000005412 00000 н
0000005434 00000 н
0000005779 00000 н
0000005802 00000 н
0000007538 00000 н
0000007561 00000 н
0000009335 00000 н
0000009358 00000 н
0000011123 00000 н
0000011407 00000 н
0000011676 00000 н
0000011699 00000 н
0000013568 00000 н
0000013589 00000 н
0000013610 00000 н
0000013633 00000 н
0000015357 00000 н
0000015380 00000 н
0000016985 00000 н
0000017008 00000 н
0000018623 00000 н
0000018646 00000 н
0000020289 00000 н
0000020310 00000 н
0000020402 00000 н
0000020423 00000 н
0000020515 00000 н
0000020537 00000 н
0000021600 00000 н
0000021622 00000 н
0000022152 00000 н
0000022174 00000 н
0000023284 00000 н
0000023307 00000 н
0000025921 00000 н
0000025943 00000 н
0000026829 00000 н
0000026852 00000 н
0000030807 00000 н
0000030830 00000 н
0000033560 00000 н
0000033583 00000 н
0000037208 00000 н
0000037231 00000 н
0000042821 00000 н
0000042844 00000 н
0000047500 00000 н
0000047523 00000 н
0000052914 00000 н
0000052937 00000 н
0000058591 00000 н
0000058614 00000 н
0000063923 00000 н
0000063946 00000 н
0000068651 00000 н
0000068674 00000 н
0000074485 00000 н
0000074508 00000 н
0000080156 00000 н
0000080179 00000 н
0000084762 00000 н
0000084785 00000 н
0000089916 00000 н
0000089939 00000 н
0000094739 00000 н
0000094762 00000 н
0000099794 00000 н
0000099817 00000 н
0000104530 00000 н
0000104553 00000 н
0000110328 00000 н
0000110351 00000 н
0000114046 00000 н
0000114069 00000 н
0000119022 00000 н
0000119045 00000 н
0000123331 00000 н
0000123354 00000 н
0000129428 00000 н
0000129451 00000 н
0000134783 00000 н
0000134806 00000 н
0000140299 00000 н
0000002477 00000 н
0000002897 00000 н
трейлер
]
>>
startxref
0
%%EOF
696 0 объект
>
эндообъект
697 0 объект
>
эндообъект
790 0 объект
>
ручей
Hb««\ À
(PDF) Токсикокинетика и токсикодинамика алкалоидов аяуаски N,N-диметилтриптамин (ДМТ), гармин, гармалин и тетрагидрогармин: клиническое и судебно-медицинское воздействие
Pharmaceuticals 2020,13, 334 34 из 36
5 191.
Гомес, Н.Г.; Кампос, М.Г.; Orf
ã
o, J.M.; Рибейро, К.А. Растения с нейробиологической активностью как потенциальные мишени
для разработки лекарств. прог. Нейропсихофармакол. биол. Психиатрия 2009, 33, 1372–1389. [CrossRef]
192.
Frecska, E.; Бокор, П.; Винкельман, М. Терапевтические возможности аяуаски: возможные эффекты против различных
болезней цивилизации. Фронт. Фармакол. 2016,7, 35. [CrossRef] [PubMed]
193.
Винкельман, М. Дж. Терапевтическое применение аяуаски и других священных лекарств. В терапевтическом использовании
аяуаски; Labate, BC, Cavnar, C., Eds.; Springer: Берлин/Гейдельберг, Германия, 2014 г.; стр. 1–21.
194.
Палхано-Фонтес, Ф.; Баррето, Д.; Ониас, Х .; Андраде, KC; Новаес, М.М.; Пессоа, Дж. А.; Мота-Ролим, С.А.;
Осорио, Флорида; Санчес, Р.; Дос Сантос, Р.Г.; и другие. Быстрые антидепрессивные эффекты психоделической аяуаски
при резистентной к лечению депрессии: рандомизированное плацебо-контролируемое исследование. Психол. Мед.
Психол. Мед.
2019
,49, 655–663.
[CrossRef] [PubMed]
195.
Самойленко В.; Рахман, М.М.; Теквани, Б.Л.; Трипати, Л.М.; Ван, Ю.Х.; Хан, С.И.; Хан, И.А.;
Миллер, Л.С.; Джоши, VC; Мухаммад, I. Banisteriopsis caapi, уникальное сочетание компонентов, ингибирующих МАО
, и антиоксидантных компонентов для действия, связанного с нейродегенеративными расстройствами и болезнью Паркинсона
. Дж. Этнофармакол. 2010, 127, 357–367.[CrossRef] [PubMed]
196.
Дос Сантос, Р.Г.; Осорио, Флорида; Криппа, JAS; Халлак, J.E.C. Классические галлюциногены и нейровизуализация: систематический обзор
исследований человека: галлюциногены и нейровизуализация. Неврологи. Биоповедение.
2016
,71,
715–728. [CrossRef]
197.
Ly, C.; Греб, AC; Кэмерон, Л.П.; Вонг, Дж. М.; Барраган, Э.В.; Уилсон, ПК; Бурбах, К.Ф.; Солтанзаде
Заранди, С.; Суд, А.; Пэдди, М.Р.; и другие. Психоделики способствуют структурной и функциональной нейронной пластичности.
Cell Rep. 2018, 23, 3170–3182. [CrossRef] [PubMed]
198.
Grob, C.S.; Маккенна, ди-джей; Каллауэй, Дж. К.; Брито, Г.С.; Невес, Э.С.; Оберлендер, Г.; Саиде, О.Л.; Лабигалини, Э.;
Такла, К.; Миранда, Коннектикут; и другие. Человеческая психофармакология хоаски, растительного галлюциногена, используемого в ритуальном контексте
в Бразилии. Дж. Нерв. Мент. Дис. 1996, 184, 86–94. [CrossRef] [PubMed]
199.Гроб, К. С. Психология аяуаски. В Аяхуаске: галлюциногены, сознание и дух природы;
Мецнер, Р., изд.; Thunder’s Mouth Press: New York, NY, USA, 1999.
200.
Da Silveira, DX; Гроб, CS; де Риос, доктор медицины; Лопес, Э.; Алонсо, Л.К.; Такла, К.; Doering-Silveira, E. Ayahuasca
в подростковом возрасте: предварительная психиатрическая оценка. Дж. Психоакт. Наркотики 2005, 37, 129–133. [CrossRef]
201.
Деринг-Сильвейра, Э.; Лопес, Э.; Гроб, CS; де Риос, доктор медицины; Алонсо, Л.К.; Такла, К.; Ширакава, И. ; Бертолуччи, П. Х.;
; Бертолуччи, П. Х.;
Да Силвейра, Д.Х. Аяхуаска в подростковом возрасте: нейропсихологическая оценка. Дж. Психоакт. Наркотики
2005
,37,
123–128. [CrossRef] [PubMed]
202.
Hendricks, P.S.; Торн, CB; Кларк, CB; Кумбс, Д.В.; Johnson, M.W. Использование классических психоделиков связано с уменьшением психологического стресса и суицидальных наклонностей у взрослого населения Соединенных Штатов.Дж. Психофармакол.
2015, 29, 280–288. [CrossRef]
203.
Lawn, W.; Халлак, Дж. Э.; Криппа, Дж. А.; Дос Сантос, Р.; Порфи, Л.; Барратт, М.Дж.; Феррис, Дж. А.; Уинсток, А.Р.;
Морган, C.J.A. Самочувствие, проблематичное потребление алкоголя и острые субъективные эффекты наркотиков у
потребителей аяхуаски за последний год: большой международный онлайн-опрос с самостоятельным отбором. науч. Rep. 2017,7, 15201. [CrossRef]
204.
Frecska, E.; Сабо, А .; Винкельман, М.Дж.; Луна, LE; МакКенна, Д.Дж. Возможно, рецептор сигма-1 опосредует
роль диметилтриптамина в защите тканей, регенерации и иммунитете. Дж. Нейрал. Трансм.
Дж. Нейрал. Трансм.
2013
,120,
1295–1303. [CrossRef] [PubMed]
205.
Ruscher, K.; Wieloch, T. Участие рецептора сигма-1 в нейродегенерации и нейровосстановлении.
J. Pharmacol. науч. 2015, 127, 30–35. [CrossRef]
206.
Фарзин Д.; Мансури, Н.Антидепрессантоподобный эффект гармана и других бета-карболинов в тесте принудительного плавания у мышей
. Евро. Нейропсихофармакол. 2006, 16, 324–328. [CrossRef]
207.
Дай, Ф.; Чен, Ю .; Песня, Ю.; Хуанг, Л.; Чжай, Д.; Донг, Ю .; Лай, Л.; Чжан, Т .; Ли, Д.; Панг, X .; и другие. Природный
низкомолекулярный гармин ингибирует ангиогенез и подавляет рост опухоли за счет активации p53 в
эндотелиальных клетках. PLoS ONE 2012,7, e52162. [Перекрестная ссылка]
208.
Реус, Г.З.; Стрингари, РБ; де Соуза, Б.; Петронильо, Ф.; Дал-Пиццол, Ф.; Халлак, Дж. Э.; Зуарди, А.В.; Криппа, Дж. А.;
Quevedo, J. Хармин и имипрамин способствуют антиоксидантной активности в префронтальной коре и гиппокампе.
Оксид. Мед. Клетка. Лонгев. 2010, 3, 325–331. [CrossRef] [PubMed]
209.
Качборян-Нето, А.; Сантос, WT; Никасио, К.Дж.; Корреа, JOA; Мургу, М .; Мартинс, ТММ; Гомес, Д.А.;
Гус, AM; Соареш, М.Г.; Диас, Д.Ф.; и другие. Нейропротекторный потенциал аяуаски и нецелевые метаболические анализы
: применимость к болезни Паркинсона. Дж. Этнофармакол.
2020
,255, 112743. [CrossRef]
[PubMed]
Влияние ингибирующего роста Peganumharmala L. на две клеточные линии рака молочной железы
ВВЕДЕНИЕ 90
Веками лекарственные травы использовались для лечения многих различных заболеваний, и в настоящее время им уделяется повышенное внимание, поскольку они дешевы, доступны в местном масштабе и имеют меньше побочных эффектов.В исследовании подсчитано, что 60-80% антибактериальных и противораковых препаратов были получены из натуральных продуктов [1].
Пеганум гармала L . (род из семейства Nitrariaceae ), также известный как сирийская рута, представляет собой лекарственное растение, распространенное в полузасушливых районах Северо-Западной Индии, Северной Африки и Центральной Азии. Это растение известно как «Эспанд» в Иране, «Хармель» в Северной Африке и «Африканская рута», «Мексиканская рута» или «Турецкая рута» в Соединенных Штатах [2]. Период цветения с марта по апрель.Плоды представляют собой шаровидные коробочки с 3 камерами, содержащие многочисленные мелкие темно-коричневые семена длиной 3-4 мм [3]. Плоды используются в качестве обезболивающего и антисептического средства в народной медицине [3], а недавние фармацевтические исследования доказали антибактериальные и антипротозойные свойства P.harmala [4,5].
Это растение известно как «Эспанд» в Иране, «Хармель» в Северной Африке и «Африканская рута», «Мексиканская рута» или «Турецкая рута» в Соединенных Штатах [2]. Период цветения с марта по апрель.Плоды представляют собой шаровидные коробочки с 3 камерами, содержащие многочисленные мелкие темно-коричневые семена длиной 3-4 мм [3]. Плоды используются в качестве обезболивающего и антисептического средства в народной медицине [3], а недавние фармацевтические исследования доказали антибактериальные и антипротозойные свойства P.harmala [4,5].
Сообщалось, что это растение обладает противоопухолевым действием, сосудорасширяющим действием, анти-ВИЧ, антиоксидантной активностью, иммуномодулирующими свойствами и гипогликемическим действием [3,6]. Выявлено, что некоторые фармакологические эффекты P.гармала может быть отнесена к его β-карболиновым алкалоидам и производным хиназолина [3]. Алкалоиды, флавоноиды и антрахиноны являются основными фитохимическими соединениями P. harmala .
harmala .
β-карболин, как и гармалин , гармин , гармалол , гармол и тетрагидро гармин , являются основными алкалоидами, присутствующими в P.harmala . Херрайс и др. (2010) определили, что разные части растения содержат разный процент вышеупомянутых алкалоидов, например, семена и корни содержат самые высокие уровни алкалоидов, однако стебли и листья содержат меньшее количество, а цветы не содержат алкалоидов.Herraiz подтвердил, что гармин и гармалин накапливаются в сухих семенах в количестве 4,3% и 5,6% (вес/вес) соответственно, гармалол в количестве 0,6% и тетрагидро гармин в количестве 0,1% (вес/вес) и корнях. содержали гармина и гармола с содержанием 2,0% и 1,4% (мас./мас.) соответственно [7]. Harmaline (дигидро-β-карболиновый алкалоид) и гармин (полноценный ароматический β-карболиновый алкалоид) являются основными алкалоидами, присутствующими в семенах и корнях P. гармала L. [8], ингибируют моноаминоксидазу А (МАО) как основной инактиватор моноаминергических нейротрансмиттеров, ответственный за ряд неврологических расстройств [7,9]. Кроме того, гармин показал значительное ингибирование опухоли у мышей с раком легких Льюис, саркомой 180 или опухолью HepA [10].
гармала L. [8], ингибируют моноаминоксидазу А (МАО) как основной инактиватор моноаминергических нейротрансмиттеров, ответственный за ряд неврологических расстройств [7,9]. Кроме того, гармин показал значительное ингибирование опухоли у мышей с раком легких Льюис, саркомой 180 или опухолью HepA [10].
Рак как одна из основных проблем общественного здравоохранения в мире является второй по значимости причиной смерти после сердечно-сосудистых заболеваний [11]. В экономически развитых и развивающихся странах рак молочной железы у женщин и рак легкого у мужчин являются наиболее часто диагностируемыми онкологическими заболеваниями [12].Рак молочной железы представляет собой сложное и гетерогенное заболевание с генетическими и экологическими факторами риска. Заболеваемость и смертность от этого рака растут во многих странах Африки и Азии [12], поэтому изучение рака молочной железы имеет решающее значение для значительного улучшения здоровья во всем мире.
Клеточные линии рака молочной железы классифицируются на основе гистологического типа, степени опухоли, статуса лимфатических узлов и наличия прогностических маркеров, таких как рецептор эстрогена (ER) и рецептор 2 эпидермального фактора роста человека (HER2) [13]. Клеточные линии рака молочной железы, такие как MDA-MB-231, SKBR-3, MCF-12A, HBL101, MDA-MD-435, MCF-7, HS598T [14], часто используются для фундаментальных исследований рака. Ингибирующее рост действие плодов T erminalia chebula было подтверждено Saleem et al. на нескольких линиях злокачественных клеток, включая линии MCF7, S115, HOS-1, PC-3 и PNT1A. (2002) [15]. Рива и др. (2001) изучали антипролиферативное действие экстрактов Uncaria tomentosa и его фракций на рост линии клеток рака молочной железы MCF7 [16].Хостанска и др. (2004) изучали антипролиферативную активность экстрактов C. racemosa (изопропанольный и этанольный) на клетках рака молочной железы с положительным рецептором эстрогена MCF7 и отрицательным по рецептору эстрогена MDA-MB231 с помощью анализа WST-1 [17]. Положительные эффекты водных экстрактов 12 китайских лекарственных трав, Anemarrhena Asphodeloides , Artemisia Argyi , Commiphora Myrrha , Gleditsnea Inde , Gleditsia Sinensis , Lueustrum Lucidum , Rhum Cordifolia , Salvia chinensis , Scutellaria barbata , Uncaria rhychophylla и Vaccaria segetalis были оценены на предмет их антипролиферативной активности на восьми линиях раковых клеток Shoemaker et al.
Клеточные линии рака молочной железы, такие как MDA-MB-231, SKBR-3, MCF-12A, HBL101, MDA-MD-435, MCF-7, HS598T [14], часто используются для фундаментальных исследований рака. Ингибирующее рост действие плодов T erminalia chebula было подтверждено Saleem et al. на нескольких линиях злокачественных клеток, включая линии MCF7, S115, HOS-1, PC-3 и PNT1A. (2002) [15]. Рива и др. (2001) изучали антипролиферативное действие экстрактов Uncaria tomentosa и его фракций на рост линии клеток рака молочной железы MCF7 [16].Хостанска и др. (2004) изучали антипролиферативную активность экстрактов C. racemosa (изопропанольный и этанольный) на клетках рака молочной железы с положительным рецептором эстрогена MCF7 и отрицательным по рецептору эстрогена MDA-MB231 с помощью анализа WST-1 [17]. Положительные эффекты водных экстрактов 12 китайских лекарственных трав, Anemarrhena Asphodeloides , Artemisia Argyi , Commiphora Myrrha , Gleditsnea Inde , Gleditsia Sinensis , Lueustrum Lucidum , Rhum Cordifolia , Salvia chinensis , Scutellaria barbata , Uncaria rhychophylla и Vaccaria segetalis были оценены на предмет их антипролиферативной активности на восьми линиях раковых клеток Shoemaker et al. (2005)[18]. Кроме того, алкалоиды, такие как винбластин, винкристин и эллиптицин, использовались в качестве сильнодействующих противораковых средств [19]. Опухолевые клетки погибали под действием этих алкалоидов по разным механизмам, таким как индукция апоптоза, ингибирование топоизомеразы I и II [20-22].
(2005)[18]. Кроме того, алкалоиды, такие как винбластин, винкристин и эллиптицин, использовались в качестве сильнодействующих противораковых средств [19]. Опухолевые клетки погибали под действием этих алкалоидов по разным механизмам, таким как индукция апоптоза, ингибирование топоизомеразы I и II [20-22].
Алкалоиды P.harmala, , в том числе гармин и гармалин , эффективны на линии промиелоцитов человека (клетки HL60) [3]. В исследовании гармин значительно ингибировал рост опухоли у мышей с раком легких Льюис, саркомой 180 и опухолью HepA [23].На сегодняшний день не проводилось исследований для проверки прямой противоопухолевой активности экстрактов P.harmala на человека на линии клеток рака молочной железы MDA-MB-231 и Mcf-7. В этом исследовании были экстрагированы и очищены алкалоиды семян P.harmala L., содержащие активные растительные компоненты, гармин и гармалин . Для их точной идентификации использовались два метода, FT-IR и ВЭЖХ, и было исследовано их влияние на две линии раковых клеток, MDA-MB-231 и Mcf-7.
МАТЕРИАЛЫ И МЕТОДЫ
Пеганум гармала Экстракт алкалоидов
Семена P.harmal были собраны в горах Шахрекорд, провинция Чахармахало Бахтиари и Мешхед, провинция Хорасан-Разави, Иран. Шлифованное р. Семена гармалы смешивали с 50 мл ледяной уксусной кислоты (30% (вес/вес)) и перемешивали в течение 30 минут на низкой скорости. Затем смесь фильтровали через воронку Бюхнера и фильтровальную бумагу Whatman (No.5) при промывании 20 мл ледяной уксусной кислоты (30% (вес/вес)) еще раз.
Фильтрат трижды промывали петролейным эфиром:этилацетатом (1:1) для удаления органических примесей в делительной воронке. Водный слой собирали и обильно добавляли 10 М гидроксида натрия до получения мутного вида. Органическую часть, содержащую в основном алкалоиды , гамалин и гармин , улавливали в хлороформной фазе (100 мл x три раза) в делительной воронке.Наконец, растворитель удаляли с помощью роторного испарителя.
ВЭЖХ-анализ
ВЭЖХ-анализ был выполнен с использованием серии Cecil 1100 (Cecil Inst., Ltd., Кембридж, Соединенное Королевство), оснащенной насосом серии 1100, детектором УФ-поглощения и колоночной печью (CTS-30 Younglin, Корея) для обнаружения цикломальтооктадекаозы (CD18). ), циклические олигосахариды, состоящие из 18 звеньев D-глюкозы. Подвижная фаза состояла из калий-фосфатного буфера (10 мМ рН 7) и ацетонитрила (50:50 по объему) со скоростью потока 1.5 мл.мин -1 при комнатной температуре (25°С).
Индивидуальные исходные растворы гармина и гармалина (Sigma, США) готовили в пяти концентрациях 100-1000 мкг.мл -1 в метаноле и использовали для построения стандартной кривой.
Инфракрасная спектроскопия с преобразованием Фурье (FTIR)
Спектры ИК-Фурье-спектроскопии записывали на ИК-Фурье-спектрофотометре (BRUKER, Германия) с использованием дисков KBr. П.Экстракты гармалы (2 мкл) наносили на диски KBr с образованием тонких жидких пленок для инфракрасного спектрометрического анализа. Диски имели диаметр около 5 мм и толщину 1 мм. Прибор работал при продувке сухим воздухом, и сканы были получены при скорости сканирования 2 мм.с -1 с разрешением 4 см -1 в области 4000-400 см -1 .
Диски имели диаметр около 5 мм и толщину 1 мм. Прибор работал при продувке сухим воздухом, и сканы были получены при скорости сканирования 2 мм.с -1 с разрешением 4 см -1 в области 4000-400 см -1 .
Линии клеток
Две клеточные линии карциномы молочной железы человека MDA-MB-231 и Mcf-7 были пожертвованы Dr.Моса Гардане и инж. Азин Голами (NIGEB). Клеточная линия MDA-MB-231 является отрицательной для ER, PR и HER2 и положительной для EGFR, однако клеточная линия Mcf-7 положительна для рецептора эстрогена (ER) [13]. Линии раковых клеток культивировали в среде DMEM с 100 ед.мл -1 пенициллина, 100 мкг.мл -1 стрептомицина и 10% эмбриональной бычьей сыворотки (FBS). Клетки выращивали при 37°С во влажном инкубаторе с 5% CO 2 .
Химикаты
3-(4,5-Диметилтиазол-2-ил)-2,5-дифенилтетразолия бромид (МТТ), гармин (286044-1G), гармалин (51330-1G) были приобретены у Sigma; DMEM, трипсин и FBS были приобретены у Invitrogen-Life Technologies.
Жизнеспособность клеток с помощью анализа МТТ
Жизнеспособность культивируемых клеточных линий определяли с помощью анализа МТТ, который основан на восстановлении соли тетразолия митохондриальной дегидрогеназой жизнеспособных клеток. Линии клеток высевали (8000 клеток/лунку) в 96-луночный планшет и инкубировали в течение 24 ч при 37 °C в инкубаторе с 5% CO2, затем добавляли различные концентрации экстракта алкалоидов P.harmala (1, 10, 20, 30). , 40, 50, 60, 80 и 100 мкг/мл -1 ) добавляли в каждую лунку и планшет инкубировали в течение 24, 48 и 72 часов.После инкубации в каждую лунку добавляли 20 мкл МТТ (5 мг.мл -1 ) до достижения общего реакционного объема 200 мкл. Затем планшет выдерживали в инкубаторе еще 5 часов. Затем среду обедняли и кристаллы формазана, появившиеся на последней стадии инкубации, растворяли в 200 мкл диметилсульфоксида. Процент жизнеспособных клеток определяли по различному поглощению аналитов и контролей, считываемому с помощью планшетного считывателя ELISA при 580 нм.
Статистический анализ
Статистический анализ выполнен с использованием SPSS (Статистический пакет для социальных наук).Анализ дисперсии (ANOVA) для повторных измерений и критерий Дункана были выполнены для выявления изменений среди групп. Результаты представляют собой средние значения ± стандартное отклонение по крайней мере для трех повторных определений теста МТТ. Различия с p-значениями <0,001 считались значимыми.
РЕЗУЛЬТАТЫ
Семена P.harmala были подвергнуты кислотно-щелочной экстракции для достижения содержания в них алкалоидов. Семена и корни P.harmala содержат более высокое содержание алкалоидов, в то время как листья и стебли содержат наименьшее количество, а цветки не содержат алкалоидов. Гармалин и гармин являются основными алкалоидами, присутствующими в семенах (4,3 и 5,6 % (вес/вес)) и корнях P.harmala L., однако гармалол и тетрагидрогармин составляют 0,6 и 0,1 % (мас. /мас.) [4]. Поэтому при идентификации экстракта двумя методами, ИК-Фурье и ВЭЖХ, мы регистрируем два алкалоида, h армалин и гармин по их абсолютно более высокой концентрации (рисунки 1 – 2).
/мас.) [4]. Поэтому при идентификации экстракта двумя методами, ИК-Фурье и ВЭЖХ, мы регистрируем два алкалоида, h армалин и гармин по их абсолютно более высокой концентрации (рисунки 1 – 2).
Для определения концентрации гармалина и гармина в экстракте P.гармала , калибровочные кривые были построены, как показано на рисунке 3.
Ингибирующее действие экстракта P.harmala на две клеточные линии рака молочной железы было протестировано с помощью анализа метилтиазолтетразолия (МТТ). Колориметрический анализ соли тетразолия МТТ измеряет цитотоксичность, пролиферацию клеток и активацию клеток. Уровень расщепления МТТ жизнеспособными клетками относительно связан с приращением числа клеток во времени. Было подтверждено, что количество клеток увеличивается нелинейно.Напротив, гибель клеток следует другому правилу, линейному во времени подходу во время инкубации МТТ (24).
Линии раковых клеток, MDA-MB-231 и Mcf-7, подверглись воздействию девяти концентраций экстракта P. harmala (1, 10, 20, 30, 40, 50, 60, 80 и 100 мкг/мл -1 ) в течение трех дней, чтобы оценить ингибирующее действие экстрактов на рост раковых клеток. Полумаксимальную ингибирующую концентрацию (IC 50 ) использовали для измерения подавления роста клеток для раковых линий раковых линий в различные временные интервалы 24ч, 48ч, 72ч.Поэтому было замечено, что 30 мкг.мл -1 являются конц. где 50% клеток погибают через 24 часа в клеточной линии MDA-MB-231 (рис. 4А). Та же концентрация была рассчитана для более длительного времени воздействия, приблизительно 48 и 72 часа.
harmala (1, 10, 20, 30, 40, 50, 60, 80 и 100 мкг/мл -1 ) в течение трех дней, чтобы оценить ингибирующее действие экстрактов на рост раковых клеток. Полумаксимальную ингибирующую концентрацию (IC 50 ) использовали для измерения подавления роста клеток для раковых линий раковых линий в различные временные интервалы 24ч, 48ч, 72ч.Поэтому было замечено, что 30 мкг.мл -1 являются конц. где 50% клеток погибают через 24 часа в клеточной линии MDA-MB-231 (рис. 4А). Та же концентрация была рассчитана для более длительного времени воздействия, приблизительно 48 и 72 часа.
Кроме того, ингибирующее действие экстракта на скорость роста Mcf-7 не было значительным через 24 ч даже при самой концентрированной обработке (100 мкг.мл -1 ), что не соответствует IC 50 (рис. 4B). . В этой клеточной линии кажется, что более длительное время воздействия оказывает значимое влияние на гибель клеток.40 мкг.мл -1 и 25 мкг.мл -1 были рассчитаны для IC 50 через 48 ч и 72 ч соответственно. Эти результаты подтверждают, что как концентрация, так и время воздействия оказывают значительное влияние на ингибирование роста Mcf-7 при обработке экстрактом P.harmala .
Эти результаты подтверждают, что как концентрация, так и время воздействия оказывают значительное влияние на ингибирование роста Mcf-7 при обработке экстрактом P.harmala .
Наконец, морфологические изменения клеток изучали с помощью оптической микроскопии, чтобы отличить апоптоз или некроз, ответственные за гибель клеток (рис. 5). Стрелки на фигуре 5 указывают на округлую форму клеток как в необработанных, так и в обработанных клеточных линиях MDA-MB-231 и Mcf-7 с различной концентрацией Р.экстракт гармалы , наблюдаемый под оптической микроскопией, 100X.
ОБСУЖДЕНИЕ
Травы, как природный/зеленый ресурс с разнообразным применением, включая кулинарию и лекарство, привлекали большое внимание во все века. Но в последнее время их изучают на предмет их надлежащего действия на некоторые названные заболевания [15-18]. Из них P.harmala L., известные в народной медицине, содержащие несколько алкалоидов, использовали для доказательства своего противоракового действия [7, 25 — 26]. В этом исследовании мы выделили алкалоидный экстракт семян P.harmala и идентифицировали его компонент с помощью двух аналитических методов, FTIR и ВЭЖХ. Хотя сообщается, что в экстракте P. гармала присутствуют различные алкалоиды, например, гармол , гармалол , гармин и гармалин [ 3 ], мы обнаружили гармин и гармалин в качестве основного компонента экстракта . по FTIR. Наши дальнейшие анализы с клеточными линиями обращаются к этим двум основным компонентам как к наиболее ответственным факторам таких результатов.На рис. 1С показано поглощение экстракта P. гармала по сравнению со стандартами гармин и гармалин (рис. 1 А и В) в диапазоне частот 4000 – 400 см -1 . Спектр экстракта P.harmala соответствовал стандартам harmine и harmaline , а поглощение экстракта P.harmala при разных волновых числах, 1072, 1237, 1455, 1624 и 3072, относилось к разным функциональным группам.
В этом исследовании мы выделили алкалоидный экстракт семян P.harmala и идентифицировали его компонент с помощью двух аналитических методов, FTIR и ВЭЖХ. Хотя сообщается, что в экстракте P. гармала присутствуют различные алкалоиды, например, гармол , гармалол , гармин и гармалин [ 3 ], мы обнаружили гармин и гармалин в качестве основного компонента экстракта . по FTIR. Наши дальнейшие анализы с клеточными линиями обращаются к этим двум основным компонентам как к наиболее ответственным факторам таких результатов.На рис. 1С показано поглощение экстракта P. гармала по сравнению со стандартами гармин и гармалин (рис. 1 А и В) в диапазоне частот 4000 – 400 см -1 . Спектр экстракта P.harmala соответствовал стандартам harmine и harmaline , а поглощение экстракта P.harmala при разных волновых числах, 1072, 1237, 1455, 1624 и 3072, относилось к разным функциональным группам. (CH), (C=O), (C=N), (OCh4), (CN) соответственно.
(CH), (C=O), (C=N), (OCh4), (CN) соответственно.
, использованный для разделения компонентов экстракта P.harmala , и сравнение результатов со стандартными растворами гармина и гармалина . Хроматограммы подтвердили присутствие гармина и гармалина , поскольку их время удерживания 8,07 и 5,32 мин соответственно соответствовало стандартному раствору (рис. 2А, В и С).
Дальнейший анализ ВЭЖХ показал, что гармалин имеет более высокую концентрацию в P.Экстракт гармалы . Посредством построения калибровочных кривых были указаны концентрации 640 мкг.мл -1 и 189 мкг.мл -1 для гармалина и гармина, соответственно.
Сообщается, что гармин и гармалин имеют сходное фармацевтически эквивалентное действие; однако гармин считается менее ядовитым; поэтому его более выгодно использовать в терапии рака. В настоящем исследовании мы использовали всего P. экстракт алкалоидов гармалы , чтобы показать его эффективность против роста двух клеточных линий рака молочной железы, in vitro . Результаты анализа степени ингибирования пролиферации (рис. 4А и В) подтвердили, что чем более концентрированный экстракт мы использовали, тем больше достигался потенциал ингибирования роста клеток. Однако время воздействия, как еще один параметр, играет важную роль в повышении эффективности подавления роста клеток. Хотя нам пришлось заключить этот параметр как функцию других параметров, поскольку время воздействия не давало существенной разницы при обработке MDA-MB-231 P.гармала , так как подавление роста клеток для линии рака MDA-MB-231 происходило за меньшее время (24 часа) по сравнению с Mcf-7. Напротив, время воздействия, по-видимому, имеет решающее значение в случае обработки Mcf-7 экстрактом P.harmala , при более длительном времени воздействия более сильно снижается скорость роста клеток, особенно через 72 часа. Учитывая 30-40 мкг.
экстракт алкалоидов гармалы , чтобы показать его эффективность против роста двух клеточных линий рака молочной железы, in vitro . Результаты анализа степени ингибирования пролиферации (рис. 4А и В) подтвердили, что чем более концентрированный экстракт мы использовали, тем больше достигался потенциал ингибирования роста клеток. Однако время воздействия, как еще один параметр, играет важную роль в повышении эффективности подавления роста клеток. Хотя нам пришлось заключить этот параметр как функцию других параметров, поскольку время воздействия не давало существенной разницы при обработке MDA-MB-231 P.гармала , так как подавление роста клеток для линии рака MDA-MB-231 происходило за меньшее время (24 часа) по сравнению с Mcf-7. Напротив, время воздействия, по-видимому, имеет решающее значение в случае обработки Mcf-7 экстрактом P.harmala , при более длительном времени воздействия более сильно снижается скорость роста клеток, особенно через 72 часа. Учитывая 30-40 мкг. мл -1 экстракта как IC 50 , можно сделать вывод, что концентрация экстракта имеет большее значение, чем время воздействия, для обеспечения более высокого цитотоксического эффекта и гибели клеток.Кроме того, стоит отметить, что клеточная линия MDA-MB-231 естественным образом имеет более высокую скорость роста, чем клеточная линия Mcf-7, что может объяснить более высокое ингибирование роста клеточной линии MDA-MB-231 экстрактом P.harmala . В других исследованиях, посвященных лечению клеточной линии рака молочной железы, использовалась заместительная гормональная терапия. Хостанска и др. 2004 обнаружили, что пролиферативная активность и гибель клеток возникают при воздействии MCF-7 и MDA-MB231 на изопропанольные и этанольные экстракты Cimicifuga racemosa [17].Эффективная доза для IC 50 составила 80,6±17,7 мкг.мл -1 в клетках MCF-7 и 58,6±12,6 мкг.мл -1 в клетках MDA-MB231 при использовании этанольного экстракта.
мл -1 экстракта как IC 50 , можно сделать вывод, что концентрация экстракта имеет большее значение, чем время воздействия, для обеспечения более высокого цитотоксического эффекта и гибели клеток.Кроме того, стоит отметить, что клеточная линия MDA-MB-231 естественным образом имеет более высокую скорость роста, чем клеточная линия Mcf-7, что может объяснить более высокое ингибирование роста клеточной линии MDA-MB-231 экстрактом P.harmala . В других исследованиях, посвященных лечению клеточной линии рака молочной железы, использовалась заместительная гормональная терапия. Хостанска и др. 2004 обнаружили, что пролиферативная активность и гибель клеток возникают при воздействии MCF-7 и MDA-MB231 на изопропанольные и этанольные экстракты Cimicifuga racemosa [17].Эффективная доза для IC 50 составила 80,6±17,7 мкг.мл -1 в клетках MCF-7 и 58,6±12,6 мкг.мл -1 в клетках MDA-MB231 при использовании этанольного экстракта. Их результаты подтвердили апоптозный способ гибели клеток согласно микроскопическому исследованию и дальнейшему анализу.
Их результаты подтвердили апоптозный способ гибели клеток согласно микроскопическому исследованию и дальнейшему анализу.
Более того, морфологические изменения формы клеток, исследованные с помощью оптической микроскопии, показали, что летальное действие экстракта P.harmala на линию раковых клеток могло быть своего рода гибелью клеток, апоптозом, так как не наблюдалось пермеабилизации мембраны и клетки сохраняли свою мембрану. нетронутый.
Однако мы считаем, что для подтверждения этого наблюдения необходимо следить за другими параметрами апоптоза на молекулярном уровне (активация каспазы , высвобождение цитохрома с и фрагментация олигонуклеосомной ДНК). Сходные результаты Zhao and Wink, 2013 показывают, что гармин индуцирует процесс старения в клетке, который приводит к ускоренной гибели клеток [27] . Кроме того, наш диагноз гибели клеток апоптоза подтверждает Hostanska et al. 2004 [17].
В заключение, результаты текущего исследования касаются противоракового эффекта P. гармала L. на его алкалоидные компоненты в основном хамин и гармалин . Воздействие экстракта P.harmala против двух линий раковых клеток, MDA-MB-231 и Mcf-7, показало ингибирование роста клеток, а при более высокой концентрации/более длительном времени происходила полная гибель клеток. Уровень смертности клеток и данные IC 50 подтвердили зависимое от дозы/времени ингибирующее действие P.harmala на эту линию раковых клеток.
гармала L. на его алкалоидные компоненты в основном хамин и гармалин . Воздействие экстракта P.harmala против двух линий раковых клеток, MDA-MB-231 и Mcf-7, показало ингибирование роста клеток, а при более высокой концентрации/более длительном времени происходила полная гибель клеток. Уровень смертности клеток и данные IC 50 подтвердили зависимое от дозы/времени ингибирующее действие P.harmala на эту линию раковых клеток.
Предлагается провести дальнейшие исследования для выяснения механизма действия как гармина , так и гармалина на большем количестве линий раковых клеток человека, и возможное использование этих растительных соединений активного начала в будущих противораковых фармацевтических препаратах является значительным.
Благодарности
Нет подтверждения.
Вклад автора
Все авторы в равной степени участвовали в настоящем исследовании.